| Первичная моторная кора | |
| Область Бродмана 4 человеческого мозга. | |
| Первичная моторная кора показана зеленым цветом. | |
| Подробности | |
| Часть | Прецентральная извилина |
| Артерия | Передний мозг Средний мозг |
| Идентификаторы | |
| латинский | кора головного мозга моторный примус |
| NeuroNames | 1910 г. |
| НейроЛекс ID | nlx_143555 |
| FMA | 224854 |
| Анатомические термины нейроанатомии [ редактировать в Викиданных ] | |
Анимация. Первичная моторная кора ( зона Бродмана 4 ) левого полушария головного мозга показана красным.
Первичная моторная кора
( Бродманн область 4 ) является областью головного мозга , что в организме человека находится в дорсальной части лобной доли . Это основная область в опорно — двигательной системе и работает совместно с другими моторными областями , включая премоторную кору , в дополнительной моторной области , задних теменной кору и несколько подкорковых областей головного мозга, планировать и выполнять движения. Первичная моторная кора анатомически определяется как область коры, содержащая крупные нейроны, известные как клетки Беца . Клетки Беца вместе с другими нейронами коры отправляют длинные аксоны вниз поспинной мозг для синапса с межнейронными цепями спинного мозга, а также непосредственно с альфа-мотонейронами в спинном мозге, которые соединяются с мышцами.
В первичной моторной коре моторное представление упорядочено (в перевернутом виде) от пальца ноги (в верхней части полушария головного мозга) до рта (внизу) вдоль складки в коре, называемой центральной бороздой.. Однако некоторые части тела могут контролироваться частично перекрывающимися областями коры. Каждое полушарие головного мозга первичной моторной коры содержит только моторное представление противоположной (контралатеральной) стороны тела. Количество первичной моторной коры, предназначенной для части тела, не пропорционально абсолютному размеру поверхности тела, а, вместо этого, относительной плотности кожных моторных рецепторов на указанной части тела. Плотность кожных моторных рецепторов на части тела обычно указывает на необходимую степень точности движения, требуемого для этой части тела. По этой причине человеческие руки и лицо имеют гораздо большее изображение, чем ноги.
Об открытии первичной моторной коры и ее взаимосвязи с другими моторными кортикальными областями см. Основную статью о моторной коре .
СОДЕРЖАНИЕ
- 1 Структура 1.1 Путь
- 1.2 Кортикомоторнейроны
- 1.3 Кровоснабжение
- 2.1 Гомункул
Структура [ править ]
Первичная моторная кора человека расположена на передней стенке центральной борозды. Он также частично выходит вперед из борозды на прецентральную извилину. Спереди первичная моторная кора ограничена набором областей, которые лежат на прецентральной извилине и которые обычно считаются составляющими латеральной премоторной коры. Сзади первичная моторная кора ограничена первичной соматосенсорной корой, которая лежит на задней стенке центральной борозды. Вентрально первичная моторная кора ограничена корой островка в боковой борозде. Первичная моторная кора простирается дорсально до верхней части полушария, а затем продолжается до медиальной стенки полушария.
Расположение первичной моторной коры наиболее очевидно при гистологическом исследовании из-за наличия характерных клеток Беца . Слой V первичной моторной коры содержит гигантские (70-100 мкм ) пирамидные нейроны, которые являются клетками Беца. Эти нейроны посылают длинные аксоны к контрлатеральным моторным ядрам черепных нервов и к нижним мотонейронам вентрального рога спинного мозга . Эти аксоны являются частью кортикоспинального тракта.. Клетки Беца составляют лишь небольшой процент кортикоспинального тракта. По некоторым оценкам, они составляют около 10% нейронов первичной моторной коры, проецируемых в спинной мозг [1], или около 2-3% общей корковой проекции в спинной мозг. [2] Хотя клетки Беца не составляют весь моторный выход коры, они, тем не менее, являются четким маркером для первичной моторной коры. Эта область коры, характеризующаяся наличием клеток Беца, была названа Бродманном областью 4.
Путь [ править ]
По мере того, как первичные моторные аксоны проходят вниз через белое вещество головного мозга , они сближаются и образуют часть задней
части внутренней капсулы .
Они продолжаются вниз в ствол мозга , где некоторые из них, переходя на контралатеральную сторону, распространяются на двигательные ядра черепных нервов . ( Примечание
: несколько синапсов моторных волокон с нижними мотонейронами на той же стороне ствола мозга ).
После перехода на контралатеральную сторону в продолговатом мозге ( пирамидный перекрест ) аксоны перемещаются по спинному мозгу по боковому кортикоспинальному тракту
.
Волокна, которые не пересекаются в стволе головного мозга, проходят по отдельному вентральному кортикоспинальному тракту , и большинство из них переходят на контралатеральную сторону спинного мозга незадолго до достижения нижних мотонейронов . Помимо основного кортикоспинального тракта, моторная кора проецируется на другие корковые и подкорковые области, включая полосатое тело, гипоталамус, средний и задний мозг, а также базальные ганглии таламуса, средний мозг и продолговатый мозг [3]
Кортикомоторнейроны [ править ]
Кортикомоторнейроны
— это нейроны первичной коры, которые проецируются непосредственно на мотонейроны вентрального рога спинного мозга. [4] [5] Аксоны кортико-моторных нейронов оканчиваются на спинномозговых мотонейронах нескольких мышц, а также на спинномозговых интернейронах. [4] [5] Они уникальны для приматов, и было высказано предположение, что их функция заключается в адаптивном управлении дистальными отделами конечностей (например, руками), включая относительно независимое управление отдельными пальцами. [5] Кортикомоторнейроны до сих пор были обнаружены только в первичной моторной коре, но не во вторичных моторных областях. [5]
Кровоснабжение [ править ]
Ветви средней мозговой артерии обеспечивают большую часть артериального кровоснабжения первичной моторной коры .
Медиальная сторона (области ног) снабжается ветвями передней мозговой артерии .
Субрегионы [ править ]
По крайней мере, шесть областей теперь распознаются в более крупном регионе, когда-то определяемом как SMA. Эти подразделения наиболее подробно изучены в мозге обезьян. Самая передняя часть в настоящее время обычно называется пре-SMA. [1] [2] [3] Он имеет редкие связи со спинным мозгом или первичной моторной корой или вообще не имеет их, а также имеет обширные связи с префронтальными областями. [1] [4] [5] [6] [7]
Поле дополнительных глаз (SEF) является относительно передней частью SMA , что, при стимуляции, вызывает движения головы и глаз и , возможно , движения конечностей и туловища. [8] [9] [10] [11]
Дам и Стрик [5] выдвинули гипотезу на основе цитоархитектуры и связей со спинным мозгом, что часть SMA в поясной борозде в медиальной части полушария может быть разделена на три отдельные области — поясные двигательные области. Функции двигательных зон поясной извилины систематически не изучены.
Собственно SMA у обезьян теперь ограничивается областью на макушке полушария и частично распространяется на медиальную стенку, непосредственно перед представлением первичной двигательной ноги. Собственно SMA проецируется непосредственно на спинной мозг и, следовательно, является одной из основных выходных областей корковой моторной системы. [5] [12] [13] [14] [15] [16]
Недавно Zhang et al. [17] исследовали функциональные подразделения медиальной SFC на основе связи всего мозга, охарактеризованной на основе большого набора данных фМРТ в состоянии покоя. Помимо воспроизведения границ между SMA и preSMA, текущие результаты подтверждают функциональную разницу между задним и передним пре-SMA. В отличие от задней пре-SMA, передняя пре-SMA связана с большей частью префронтальных, но не сомато-моторных областей. В целом, SMA прочно связана с таламусом и эпиталамусом , задняя пре-SMA — с скорлупой , паллидумом и STN, а передняя пре-SMA — с хвостатым ядром , причем хвостатое ядро демонстрирует значительную асимметрию полушария.
Функция [ править ]
Гомункул [ править ]
Различные части тела в первичной моторной коре широко представлены в структуре, называемой моторным гомункулом (латинское: маленький человек
). [6] Область ног расположена близко к средней линии, во внутренних частях моторной области, переходящей в медиальную продольную щель . Боковая, выпуклая сторона первичной моторной коры расположена сверху вниз в областях, которые соответствуют ягодицам, торсу, плечу, локтю, запястью, пальцам, большим пальцам, векам, губам и челюсти. Моторная область руки и кисти является самой большой и занимает часть прецентральной извилины между ногой и областью лица.
Эти области не пропорциональны их размеру на теле, при этом губы, части лица и руки представлены особенно большими участками. После ампутации или паралича моторные области могут сместиться, чтобы приспособиться к новым частям тела.
Нейронный сигнал от таламуса [ править ]
Первичная моторная кора получает таламические входы от различных ядер таламуса. Среди прочего:
— Вентрально-латеральное ядро афферентов мозжечка
— Вентральное переднее ядро афферентов базальных ганглиев
Альтернативные карты [ править ]
Карта тела в мозгу человека
Сообщалось по крайней мере о двух модификациях классического соматотопического упорядочения частей тела в первичной моторной коре приматов.
Во-первых, представление руки может быть организовано в виде стержня и окружения. В коре головного мозга обезьяны пальцы руки представлены в центральной области на заднем крае первичной моторной коры. Эта центральная область окружена с трех сторон (на дорсальной, передней и вентральной сторонах) более проксимальными частями руки, включая локоть и плечо. [7] [8] У людей изображение пальцев окружено дорсально, спереди и снизу изображением запястья. [9]
Вторая модификация классического соматотопического упорядочения частей тела — это двойное представление пальцев и запястья, изучаемое в основном в моторной коре головного мозга человека. Одно представление находится в задней области, называемой областью 4p, а другое — в передней области, называемой областью 4a. Задняя область может быть активирована вниманием без какой-либо сенсорной обратной связи, и было высказано предположение, что она важна для инициирования движений, в то время как передняя область зависит от сенсорной обратной связи. [10] Его также можно активировать воображаемыми движениями пальцев [11] и прослушиванием речи, не делая реальных движений. Предполагается, что эта передняя репрезентативная зона важна для выполнения движений, включающих сложные сенсомоторные взаимодействия. [12]Возможно, что область 4a у человека соответствует некоторым частям каудальной премоторной коры, как описано в коре головного мозга обезьяны.
В 2009 году сообщалось, что есть две эволюционно различных области: более старая на внешней поверхности и новая, обнаруженная в расщелине. Более старый соединяется с моторными нейронами спинного мозга через интернейроны спинного мозга. Более новый, обнаруживаемый только у обезьян и обезьян, напрямую соединяется с моторными нейронами спинного мозга. [13] Прямые связи образуются после рождения, преобладают над косвенными связями и более гибки в схемах, которые они могут развить, что позволяет постнатально обучаться сложной мелкой моторики. «Таким образом, появление« новой »области M1 во время эволюции линии приматов, вероятно, имело важное значение для повышения ловкости рук человека». [14]
Распространенные заблуждения [ править ]
Определенные заблуждения о первичной моторной коре часто встречаются во вторичных обзорах, учебниках и популярных материалах. Здесь перечислены три наиболее распространенных заблуждения.
Сегрегированная карта тела [ править ]
Одно из наиболее распространенных заблуждений о первичной моторной коре состоит в том, что карта тела четко разделена. Однако это не карта отдельных мышц или даже отдельных частей тела. Карта имеет значительное перекрытие. Это перекрытие увеличивается в более передних областях первичной моторной коры. Одна из основных целей в истории исследований моторной коры головного мозга состояла в том, чтобы определить, насколько различные части тела перекрываются или сегрегированы в моторной коре. Исследователи, которые занимались этим вопросом, обнаружили, что карта кисти, руки и плеча сильно перекрывается. [6] [8] [9] [15] [16] [17] [18] [19]Исследования, которые отображают точную функциональную связь между кортикальными нейронами и мышцами, показывают, что даже один нейрон в первичной моторной коре может влиять на активность многих мышц, связанных со многими суставами. [15] В экспериментах на кошках и обезьянах, по мере того как животные обучаются сложным, скоординированным движениям, карта в первичной моторной коре становится все более перекрывающейся, очевидно, учась интегрировать контроль над многими мышцами. [20] [21] У обезьян, когда электрическая стимуляция применяется к моторной коре в поведенческой шкале времени, она вызывает сложные, высоко интегрированные движения, такие как дотягивание рукой, имеющей форму для захвата, или поднесение руки ко рту и открывание рот. [22] [23]Этот тип свидетельств предполагает, что первичная моторная кора головного мозга, хотя и содержит приблизительную карту тела, может участвовать в интеграции мышц значимым образом, а не в разделении контроля над отдельными группами мышц. Было высказано предположение, что более глубокий принцип организации может быть картой статистических корреляций в поведенческом репертуаре, а не картой частей тела. [23] [24] В той степени, в которой репертуар движений частично разбивается на действия отдельных частей тела, карта содержит грубое и перекрывающееся расположение тела.
M1 и первичная моторная кора [ править ]
Термин «M1» и термин «первичная моторная кора» часто используются как синонимы. Однако они происходят из разных исторических традиций и относятся к разным отделам коры головного мозга. Некоторые ученые предположили, что моторная кора головного мозга может быть разделена на первичную моторную полосу, которая находится более сзади, и латеральную премоторную полосу, которая находится в более передней части. Ранние исследователи, первоначально предложившие эту точку зрения, включали Кэмпбелл, [25] Фогт и Фогт, [26] Ферстер, [27] и Фултон. [28]Другие предположили, что моторную кору нельзя разделить таким образом. Вместо этого, на этом втором изображении, так называемые первичные моторные и боковые премоторные полоски вместе составляют единую кортикальную область, называемую M1. Вторая моторная зона на медиальной стенке полушария была названа М2 или дополнительной моторной зоной . Сторонниками этой точки зрения были Пенфилд [6] и Вулси. [29] Сегодня общепринято различие между первичной моторной корой и латеральной премоторной корой. Однако термин M1 иногда ошибочно используется для обозначения первичной моторной коры. Строго говоря, M1 относится к единой карте, которая, по мнению некоторых предыдущих исследователей, охватывала как первичную моторную, так и латеральную премоторную кору.
Клетки Беца как последний общий путь [ править ]
В клетки Беца , или гигантские пирамидальные клетки в первичной моторной коре, иногда ошибочно принимают быть единственным или основным выходом из коры головного мозга к спинному мозгу. Это старая ошибка, относящаяся, по крайней мере, к Кэмпбеллу в 1905 году. [25] Тем не менее, клетки Бец составляют лишь около 2-3% нейронов, которые проецируются из коры в спинной мозг [2], и только около 10% нейронов нейроны, которые проецируются из первичной моторной коры в спинной мозг. [1] Ряд областей коры головного мозга , включая премоторную кору , дополнительную двигательную область.и даже первичная соматосенсорная кора проецируются на спинной мозг. Даже когда клетки Беца повреждены, кора все еще может связываться с подкорковыми двигательными структурами и контролировать движение. Если первичная моторная кора головного мозга с ее клетками Беца повреждена, это приводит к временному параличу, и другие области коры, очевидно, могут взять на себя часть утраченной функции.
ЗОНЫ БОЛЬШИХ ПОЛУШАРИЙ
Локализация функций в больших полушариях. Кора больших полушарий головного мозга делится на основные зоны, состоящие из нескольких корковых полей. Каждая из этих зон выполняет определенную общую функцию, а составляющие ее поля специализированно участвуют в реализации отдельных элементов этой функции. Однако благодаря проводящим путям в осуществлении отдельных звеньев высшей и низшей нервной деятельности участвует несколько зон больших полушарий, определенные подкорковые центры, ядра мозгового ствола и сегменты спинного мозга.
При тонкой и точной специализации определенных групп нейронов головной и спинной мозг функционируют как единое целое. Психические функции головного мозга также не ограничены отдельными участками коры, а являются результатом совместной деятельности обширных зон больших полушарий и подкорковых центров.
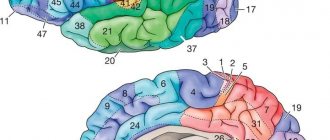
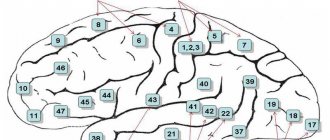
Рис. 123. Индивидуальные изменения основных полей новой коры больших полушарий у трех взрослых (А, Б, В). Цифры— поля по Бродману Моторная зона (поле 4) расположена в передней центральной извилине вдоль центральной борозды. В верхней четверти зоны находятся двигательные центры для мышц ног.
Сверху расположены нейроны, иннервирующие мышцы пальцев ног, а снизу — бедра и туловища. Две средние четверти заняты центрами для рук, выше — центр мышц лопатки, а ниже — мышц пальцев. И, наконец, в нижней четверти передней центральной извилины находятся центры мышц лица и речевого аппарата. В результате исторического развития головного мозга человека в процессе труда и речи особенно большое место занимают группы нейронов, которые вызывают сокращение мышц кисти руки, главным образом большого пальца, и мышц лица, языка и гортани. К ним поступают центростремительные волокна из проприорецепторов, входящие по задним корешкам в спинной мозг, где они поднимаются в составе заднего столба той же стороны до ядер нежного и клиновидного пучков продолговатого мозга. Из этих ядер выходят волокна вторых нейронов, образующие медиальную петлю и после перекреста достигающие ядер зрительного бугра противоположной стороны. Отсюда большая часть центростремительных волокон третьих нейронов достигает задней центральной извилины и далее поступает в переднюю центральную извилину, а меньшая часть входит в нее прямым путем. Таким образом, передняя центральная извилина посредством волокон, проходящих в проводящих путях коры, связана с задней центральной извилиной. Из моторной зоны выходят центробежные двигательные волокна пирамидных нейронов, которые составляют пирамидные проводящие пути; они достигают нейронов передних рогов спинного мозга. Моторная зона вызывает координированные движения скелетных мыщц, преимущественно на противоположной стороне тела. Она функционирует совместно с подкорковыми центрами — полосатыми телами, а также люисовым телом, красным ядром и черным веществом. При поражениях определенных участков передней центральной извилины нарушаются произвольные движения отдельных групп мышц. Неполное поражение зоны вызывает нарушение движений— парез, а полное ее разрушение — паралич. Зона кожно-мышечной чувствительности (поля 1, 2, 3, 43 и частично 5 и 7) расположена в задней центральной извилине вдоль задней центральной борозды. В этой зоне особенно сильно развиты зернистые слои коры, к которым подходят центростремительные волокна из рецепторов кожи, идущие в составе тех же проводящих путей, как и волокна из проприорецепторов. Расположение воспринимающих групп нейронов такое же, как в моторной зоне. Наибольшую поверхность занимают нейроны, воспринимающие импульсы из рецепторов кисти руки, лица, языка и гортани. Поле 7 больше других полей связано с чувствительностью руки. Зона кожно-мышечной чувствительности не полностью отграничена от моторной зоны, так как в полях 3, 4 и 5 происходит сочетание зернистых нейронов с гигантскими пирамидными нейронами. В моторной зоне находится примерно 80% двигательных нейронов, а в зоне кожно-мышечной чувствительности — 20%. В каждое полушарие поступают импульсы главным образом из рецепторов противоположной стороны тела, но также и из рецепторов той же стороны. В эту зону поступают центростремительные импульсы преимущественно из бокового и полулунного ядер зрительного бугра. При поражениях определенных участков задней центральной извилины нарушается чувствительность в отдельных участках кожи. Потеря способности узнавать предметы при их осязании обозначается как тактильная агнозия. При нарушениях функций зоны наблюдаются расстройства осязания, болевых и температурных ощущений кожи и мышечно-суставной чувствительности. Неполное поражение зоны вызывает понижение рецепции — гипостезию, а полное — ее потерю — анестезию. Лобная зона (поля 6, 5, 9, 10, 11, 44, 45, 46, 47) расположена в лобной доле впереди моторной. Она делится на премоторную и речедвигательную. Премоторная зона (поля 6, 8, 9, 10, 11) регулирует тонус скелетных мышц и координированные движения тела, ориентирующие его в пространстве. С полем 10, которое участвует в выполнении двигательных условных рефлексов, функционально связано поле 46. В премоторную зону поступают центростремительные импульсы из внутренних органов и из нее исходит значительная часть центробежных вегетативных волокон. Поэтому поражение премоторной зоны вызывает нарушение координации движений — атаксию и расстройства функций сердечнососудистой, дыхательной, пищеварительной и других систем внутренних органов. Зрительная зона (поля 17, 18, 19) расположена на внутренней поверхности затылочной доли по обеим сторонам шпорной борозды. У человека она занимает 12% общей поверхности коры. Поле 17 находится на затылочном полюсе; оно окружено полем 18, которое окружает поле 19, граничащее с задним отделом лимбической области, верхней и нижней теменными областями. В поле 17 — центральном поле зрительной зоны в 16 раз больше нейронов, чем в центральном поле слуховой зоны (поле 41), и в 10 раз больше нейронов, чем в центральном поле моторной зоны (поле 4). Это указывает на ведущее в историческом и индивидуальном развитии человека значение зрения. Из сетчатки 900 тыс.— 1 млн. центростремительных волокон зрительных нервов доходит до наружного коленчатого тела, в котором точно проецируются отдельные части сетчатки. Центростремительные волокна нейронов наружного коленчатого тела направляются в зрительную зону, преимущественно в основное зрительное поле 17. Другими промежуточными зрительными центрами, участвующими в передаче не зрительных импульсов, а глазодвигательных, являются подушка зрительного бугра и передние бугры четверохолмия. До поступления в наружное коленчатое тело волокна зрительного нерва перекрещиваются. Благодаря этому перекресту в составе зрительного пути, направляющегося в зрительную зону каждого полушария, 50% волокон своей стороны и 50% волокон противоположной стороны. В зрительную зону левого полушария поступают зрительные импульсы из левых половин сетчаток обоих глаз, а в зону правого полушария — из правых половин сетчаток обоих глаз. Поэтому разрушение одной из зрительных зон вызывает слепоту в одноименных половинах сетчаток в обоих глазах — гемианопсию. В зрительных нервах, кроме центростремительных волокон, проходят и несколько более толстые центробежные волокна к мышцам радужной оболочки и центробежные тонкие симпатические волокна из нейронов подкорковых центров. Небольшая часть центростремительных волокон зрительного нерва не прерывается в подкорковых образованиях, а прямо направляется в мозжечок и зрительные зоны больших полушарий. Разрушение обоих полей 17 вызывает полную корковую слепоту, разрушение поля 18 приводит к потере зрительной памяти при сохранении зрения, что обозначается как зрительная агнозия, а разрушение поля 19 — к потере ориентации в непривычной обстановке. Слуховая зона (поля 41, 42, 21, 22, 20, 37) расположена на поверхности височной доли, преимущественно передней поперечной височной извилины и верхней височной извилины. Поле 41, расположенное в верхней височной извилине и в передней части поперечной извилины, является проекцией кортиева органа улитки. Из органа Корти центростремительные импульсы проходят через спиральный узел по улиточному нерву, состоящему примерно из 30 тыс. волокон. В этом узле находятся первые биполярные нейроны слухового пути. Далее волокна первых нейронов передают слуховые импульсы в ядра слухового нерва в продолговатом мозге, где находятся вторые нейроны. Волокна ядер слухового нерва связываются с ядрами лицевого нерва в продолговатом мозге и глазодвигательного нерва в передних буграх среднего мозга. Поэтому при сильных звуках рефлекторно сокращаются мышцы лица, век, ушной раковины и вызываются движения глаз. Большая часть волокон ядер слухового нерва перекрещивается в варолиевом мосту, а меньшая проходит на своей стороне. Затем волокна слухового пути поступают в боковую лемнисковую петлю, которая заканчивается в задних буграх четверохолмия и во внутреннем коленчатом теле, где находятся третьи нейроны — их волокна проводят центростремительные импульсы в слуховую зону. Существуют также прямые пути, связывающие ядра слуховых нервов с мозжечком и слуховой зоной. Большая часть прямых мозжечковых путей образуется вестибулярным нервом, а меньшая— улитковым нервом, составляющими вместе общий ствол слухового нерва. Вестибулярный аппарат проецируется также в слуховой зоне. Разрушение поля 41 на одной стороне вызывает глухоту на противоположной стороне и ослабление слуха на своей стороне, а разрушение полей 41 на обеих сторонах ведет к полной корковой глухоте. Разрушение поля 22 в передней трети верхней височной извилины приводит к музыкальной глухоте — теряется восприятие интенсивности тона, тембра и ритма звуков — слуховая агнозия. Разрушение полей 21 и 20 в средней и нижней височных извилинах вызывает атаксию — расстройство равновесия и координации движений. В слуховой зоне расположен также рече-слуховой центр. Обонятельная и вкусовая зоны. Обонятельная зона находится в древней коре, в которую поступают центростремительные импульсы из обонятельных клеток. Кроме обонятельной функции, она выполняет также вкусовую и участвует в деятельности пищеварительной, выделительной и половой систем. Раньше считали, что гиппокамп выполняет обонятельную функцию. В настоящее время полагают, что вместе с лимбической системой, гипоталамической областью промежуточного мозга и гипофизом, средним и продолговатым мозгом и особенно ретикулярной формацией гиппокамп участвует в общих двигательных реакциях и вегетативных рефлексах при эмоциях. Собственно вкусовая зона, вероятно, расположена в поле 43, которое находится в нижнем отделе задней центральной извилины.
Лимбическая извилина (заднее поле 23 и переднее поле 24) и кора островка (поля 13 и 14) участвуют в высшей нервной деятельности. Все зоны коры не обособлены, а связаны между собой проводящими путями. Центры речи (поля 44, 45, 46, 39, 40, 42, 22,37). Двигательный центр речи расположен в нижней части передней центральной извилины в поле 44. У большинства правшей площадь поля 44 в левом полушарии больше, чем в правом полушарии. Поле 44 вызывает сложные сокращения речевой мускулатуры, необходимые для произнесения слов. При разрушении этого поля человек не может говорить, но может производить простейшие сокращения речевой мускулатуры — кричать и петь. Это моторная, двигательная афазия, которая в некоторых случаях проявляется в отсутствии сокращений мышц языка и остальной речевой мускулатуры. Так как в этих случаях слуховой центр речи не поврежден, то понимание речи окружающих сохраняется. При поражении поля 44 часто нарушается не только устная речь, но и внутренняя речь или способность формулировать мысли словами без их произнесения, на основе накопленных звуковых образов, имеющих определенное смысловое содержание. При этом затруднено чтение про себя, расстроена способность писать произвольно и под диктовку, но сохранено копирование букв при письме. У правшей моторная афазия наблюдается при поражении левого полушария, а левшей — правого.
Рис. 129. Локализация центров речи: 1 — двигательный, 2 — слуховой, 3 — зрительный Впереди поля 44 расположено поле 45, которое регулирует построение грамматически правильных сочетаний слов и пение. При поражении этого поля вследствие потери памяти на приемы произношения пение расстраивается. Мимика и жестикуляция, придающие речи ее выразительность, осуществляются благодаря импульсам, поступающим из поля 46 в поля 44 и 45, в поля премоторной области и в подкорковые центры. Слуховой, или сенсорный, центр речи расположен в заднем отделе левой верхней височной извилины в поле 42, которое осуществляет понимание слова при слышании его. Если поле разрушается, теряется способность понимания смысла слов, но сохраняется их восприятие как звуков — сенсорная афазия, или речевая глухота. При этом вследствие отсутствия понимания собственной речи, иногда наблюдается чрезмерная говорливость — логоррея, или словесный понос. В задней части поля 22 фиксируются связи звуковых образов слов со всеми воспринимающими зонами, в которых возникают представления о предметах и явлениях. Поэтому поражение этого поля также вызывает сенсорную афазию. Поля 39 и 40, расположенные в теменной доле рядом с полем 22, осуществляют понимание смысла сочетаний слов или фраз. Поэтому их поражение приводит к расстройству речи, которое называется семантической (смысловой) афазией. При поражении поля 39, вследствие потери способности узнавать буквы и цифры и понимать смысл видимых письменных образов слов и цифр, теряется способность читать вслух, писать и считать. Поражение поля 40 вызывает потерю способности писать, так как отсутствует ориентация движений в пространстве и нарушена их последовательность. Это отсутствие способности производить системные, целенаправленные движения (апраксия) не исключает возможности правильно совершать отдельные движения руки, не связанные с письмом. Следовательно, процесс письма у правшей осуществляется, височной, нижнетеменной и нижнелобной областями левого полушария. При поражении поля 37 вызывается потеря памяти на слова — амнестическая афазия. Таким образом, в осуществлении функции речи участвуют большие полушария головного мозга в целом, но особенная роль выполняется отдельными полями коры. У правшей в результате преимущественного развития функций правой руки и правой половины тела особенно развиты сложнейшие психические функции левого полушария головного мозга.
Похожие материалы:
Филогенез головного мозга
Строение полушарий головного мозга
Кровоснабжение головного мозга
Спинномозговая жидкость
Кодирование движения [ править ]
Эвартс [30] предположил, что каждый нейрон моторной коры вносит вклад в силу в мышце. Когда нейрон становится активным, он посылает сигнал в спинной мозг, сигнал передается на моторнейрон, моторнейрон посылает сигнал в мышцу, и мышца сокращается. Чем выше активность нейрона моторной коры, тем больше мышечная сила.
Георгопулос и его коллеги [31] [32] [33] предположили, что одна только мышечная сила — это слишком простое описание. Они обучили обезьян достигать различных направлений и отслеживали активность нейронов моторной коры. Они обнаружили, что каждый нейрон моторной коры был максимально активен в определенном направлении досягаемости и хуже реагировал на соседние направления досягаемости. На этом основании они предположили, что нейроны моторной коры, «голосуя» или объединяя свои влияния в « популяционный код », могут точно определять направление досягаемости.
Предложение о том, что нейроны моторной коры кодируют направление досягаемости, стало спорным. Скотт и Каласка [34] показали, что каждый нейрон моторной коры лучше коррелирует с деталями движения суставов и мышечной силы, чем с направлением досягаемости. Шварц и его коллеги [35] показали, что нейроны моторной коры хорошо коррелируют со скоростью руки. Стрик и его коллеги [36] обнаружили, что некоторые нейроны моторной коры активны в связи с мышечной силой, а некоторые — с пространственным направлением движения. Тодоров [37] предположил, что множество различных корреляций является результатом мышечного регулятора, в котором многие параметры движения коррелируют с мышечной силой.
Код, с помощью которого нейроны моторной коры приматов управляют спинным мозгом и, следовательно, движением, остается дискуссионным.
Определенный прогресс в понимании того, как моторная кора вызывает движение, также был достигнут в модели грызунов. Моторная кора грызунов, как и моторная кора обезьяны, может содержать подобласти, которые подчеркивают различные общие типы действий. [38] [39] Например, одна область, кажется, подчеркивает ритмический контроль взбивания . [38] [40] [41] Нейроны в этой области проецируются в определенное подкорковое ядро, в котором генератор паттернов координирует циклический ритм усов. Затем это ядро проецируется на мышцы, которые контролируют усы.
Ссылки [ править ]
- ^ a b Rivara CB, Sherwood CC, Bouras C, Hof PR (2003). «Стереологическая характеристика и пространственные модели распределения клеток Беца в первичной моторной коре головного мозга человека». Анатомическая запись, часть A: открытия в молекулярной, клеточной и эволюционной биологии
.
270
(2): 137–151. DOI : 10.1002 / ar.a.10015 . PMID 12524689 . - ^ a b Лассек AM (1941). «Пирамидальный тракт обезьяны». J. Comp.Neurol
.
74
(2): 193–202. DOI : 10.1002 / cne.900740202 . S2CID 83536088 . - «Просмотр разделов» . brainarchitecture.org
. Проверено 20 ноября 2021 . - ^ a b Сигельбаум, Стивен А .; Хадспет, AJ (2013). Принципы нейронауки
. Кандел, Эрик Р. (5-е изд.). Нью-Йорк. ISBN 9780071390118. OCLC 795553723 . - ^ a b c d Лимон, Роджер Н. (4 апреля 2008 г.). «Нисходящие пути в управлении моторикой». Ежегодный обзор нейробиологии
.
31
(1): 195–218. DOI : 10.1146 / annurev.neuro.31.060407.125547 . ISSN 0147-006X . PMID 18558853 . - ^ a b c Пенфилд У. и Болдри Э. (1937). «Соматические моторные и сенсорные представления в коре головного мозга человека при изучении с помощью электростимуляции». Мозг
.
60
(4): 389–443. DOI : 10,1093 / мозг / 60.4.389 .CS1 maint: несколько имен: список авторов ( ссылка ) - Перейти
↑ Kwan HC, MacKay WA, Murphy JT, Wong YC (1978). «Пространственная организация прецентральной коры у бодрствующих приматов. II. Двигательные выходы».
J. Neurophysiol
.
41
(5): 1120–1131. DOI : 10,1152 / jn.1978.41.5.1120 . PMID 100584 . - ^ a b Парк, М.К., Белхадж-Саиф, А., Гордон, М. и Чейни, П.Д. (2001). «Согласованные особенности в представлении передних конечностей первичной моторной коры у макак-резусов» . J. Neurosci
.
21
(8): 2784–2792. DOI : 10.1523 / JNEUROSCI.21-08-02784.2001 . PMID 11306630 . CS1 maint: несколько имен: список авторов ( ссылка ) - ^ a b Мейер, JD, Афлало, TN, Кастнер, С. и Грациано, MSA (2008). «Сложная организация первичной моторной коры человека: исследование фМРТ с высоким разрешением» . J. Neurophysiol
.
100
(4): 1800–1812. DOI : 10,1152 / jn.90531.2008 . PMC 2576195 . PMID 18684903 . CS1 maint: несколько имен: список авторов ( ссылка ) - Binkofski F, Финк GR, Гейер S, Buccino G, Грубер О, штат Нью — Джерси Шаха, Тейлор Ю.Г., Зейтц RJ, Zilles К, Freund HJ (2002). «Нервная активность в областях 4a и 4b первичной моторной коры человека по-разному модулируется вниманием к действию». J. Neurophysiol
.
88
(1): 514–519. DOI : 10,1152 / jn.2002.88.1.514 . PMID 12091573 . - Нихил Шарма; PS Джонс; Т. А. Карпентер; Жан-Клод Барон (2008). «Картирование участия BA 4a и 4p во время Motor Imagery». NeuroImage
.
41
(1): 92–99. DOI : 10.1016 / j.neuroimage.2008.02.009 . PMID 18358742 . S2CID 8673179 . - Terumitsu М, Икеда К, Kwee ИЛ и Накада, Т (2009). «Участие области 4a первичной моторной коры в сложной сенсорной обработке: исследование 3.0T fMRI». NeuroReport
.
20
(7): 679–683. DOI : 10.1097 / WNR.0b013e32832a1820 . PMID 19339906 . S2CID 23674509 . CS1 maint: несколько имен: список авторов ( ссылка ) - Rathelot, J.-A .; Стрик, Польша (20 января 2009 г.). «Подразделения первичной моторной коры на основе кортико-мотонейрональных клеток» . Proc. Natl. Акад. Sci
.
106
(3): 918–923. Bibcode : 2009PNAS..106..918R . DOI : 10.1073 / pnas.0808362106 . PMC 2621250 . PMID 19139417 . - Costandi, Mo (2009). «Эволюция ловкости рук» . Труды Национальной академии наук
.
106
(3): 918–923. DOI : 10.1073 / pnas.0808362106 . PMC 2621250 . PMID 19139417 . Проверено 29 ноября 2015 года . - ^ a b Чейни, PD & Fetz, EE (1985). «Сопоставимые паттерны облегчения мышц, вызванные отдельными кортикомотонейрональными (CM) клетками и отдельными внутрикортикальными микростимулами у приматов: доказательства функциональных групп CM-клеток». J. Neurophysiol
.
53
(3): 786–804. DOI : 10,1152 / jn.1985.53.3.786 . PMID 2984354 . - Шибер, MH & Hibbard, LS (1993). «Насколько соматотопна область моторной коры руки?». Наука
.
261
(5120): 489–492. Bibcode : 1993Sci … 261..489S . DOI : 10.1126 / science.8332915 . PMID 8332915 . - Rathelot, JA & Стрик, PL (2006). «Представление мышц в моторной коре макака: анатомическая перспектива» . Proc. Natl. Акад. Sci. США
.
103
(21): 8257–8262. Bibcode : 2006PNAS..103.8257R . DOI : 10.1073 / pnas.0602933103 . PMC 1461407 . PMID 16702556 . - Sanes, Ю.Н., Доног, ДП, Thangaraj, В., Эдельман, РР и Warach, С. (1995). «Общие нейронные субстраты, контролирующие движения рук в моторной коре человека». Наука
.
268
(5218): 1775–1777. Bibcode : 1995Sci … 268.1775S . DOI : 10.1126 / science.7792606 . PMID 7792606 . CS1 maint: несколько имен: список авторов ( ссылка ) - Доногу, ДП, Leibovic, С. и Sanes, Ю.Н. (1992). «Организация области передних конечностей в моторной коре головного мозга белки обезьяны: представление мышц пальцев, запястий и локтей». Exp. Brain Res
.
89
(1): 1–10. DOI : 10.1007 / bf00228996 . PMID 1601087 . S2CID 1398462 . CS1 maint: несколько имен: список авторов ( ссылка ) - Nudo, RJ, Милликен, GW, Дженкинс, WM и Мерцених, MM (1996). «Зависимые от использования изменения представлений о движении в первичной моторной коре взрослых беличьих обезьян» . J. Neurosci
.
16
(2): 785–807. DOI : 10.1523 / JNEUROSCI.16-02-00785.1996 . PMC 6578638 . PMID 8551360 . CS1 maint: несколько имен: список авторов ( ссылка ) - Martin, JH, Engber, Д. и Мэн, Z. (2005). «Влияние использования передних конечностей на постнатальное развитие моторного представительства передних конечностей в первичной моторной коре головного мозга кошек». J. Neurophysiol
.
93
(5): 2822–2831. DOI : 10,1152 / jn.01060.2004 . PMID 15574795 . CS1 maint: несколько имен: список авторов ( ссылка ) - Перейти
↑ Graziano, MSA , Taylor, CSR and Moore, T. (2002). «Сложные движения, вызванные микростимуляцией прецентральной коры».
Нейрон
.
34
(5): 841–851. DOI : 10.1016 / S0896-6273 (02) 00698-0 . PMID 12062029 . S2CID 3069873 . CS1 maint: несколько имен: список авторов ( ссылка ) - ^ а б Грациано, MSA (2008). Интеллектуальная машина движения
. Оксфорд, Великобритания: Издательство Оксфордского университета. - Грациано, MSA и Эфлало, TN (2007). «Отображение поведенческого репертуара на кору» . Нейрон
.
56
(2): 239–251. DOI : 10.1016 / j.neuron.2007.09.013 . PMID 17964243 . CS1 maint: несколько имен: список авторов ( ссылка ) - ^ a b Кэмпбелл, AW (1905). Гистологические исследования по локализации церебральной функции . Кембридж, Массачусетс: Издательство Кембриджского университета.
- Перейти
↑ Vogt, C. and Vogt, O. (1919). «Ergebnisse unserer Hirnforschung».
Journal für Psychologie und Neurologie
.
25
: 277–462.CS1 maint: несколько имен: список авторов ( ссылка ) - Перейти
↑ Foerster, O (1936). «Моторная кора человека в свете доктрин Хьюлингса Джексона».
Мозг
.
59
(2): 135–159. DOI : 10,1093 / мозг / 59.2.135 . - Перейти
↑ Fulton, J (1935). «Примечание к определению« моторной »и« премоторной »областей».
Мозг
.
58
(2): 311–316. DOI : 10,1093 / мозг / 58.2.311 . - ↑
Woolsey, CN, Settlage, PH, Meyer, DR, Sencer, W., Hamuy, TP и Travis, AM (1952). «Паттерн локализации в премоторных областях и их связь с понятием премоторной зоны».
Ассоциация исследований нервных и психических заболеваний
. Нью-Йорк, Нью-Йорк: Raven Press.
30
: 238–264.CS1 maint: несколько имен: список авторов ( ссылка ) - Evarts, Е. В. (1968). «Связь активности пирамидного тракта с силой, прилагаемой при произвольном движении». J. Neurophysiol
.
31
(1): 14–27. DOI : 10,1152 / jn.1968.31.1.14 . PMID 4966614 . - Георгопулоса, AP, Kalaska, JF, Caminiti, Р. Масси, JT (1982). «О связи между направлением двумерных движений руки и разрядом клеток в моторной коре приматов» . J. Neurosci
.
2
(11): 1527–1537. DOI : 10.1523 / JNEUROSCI.02-11-01527.1982 . PMC 6564361 . PMID 7143039 . CS1 maint: несколько имен: список авторов ( ссылка ) - Георгопулоса А.П., Кеттнер, RE и Шварц, А. Б. (1988). «Моторная кора приматов и движения свободной руки к визуальным целям в трехмерном пространстве. II. Кодирование направления движения нейрональной популяцией» . J. Neurosci
.
8
(8): 2928–2937. DOI : 10.1523 / JNEUROSCI.08-08-02928.1988 . PMC 6569382 . PMID 3411362 . CS1 maint: несколько имен: список авторов ( ссылка ) - Георгопулоса А.П., Шварц, AB и Кеттнер, RE (1986). «Нейронное популяционное кодирование направления движения». Наука
.
233
(4771): 1416–1419. Bibcode : 1986Sci … 233.1416G . DOI : 10.1126 / science.3749885 . PMID 3749885 . CS1 maint: несколько имен: список авторов ( ссылка ) - Scott, SH & Kalaska, JF (1995). «Изменения активности моторной коры во время движений по достижению схожей траектории движения рук, но с разным положением рук». J. Neurophysiol
.
73
(6): 2563–2567. DOI : 10,1152 / jn.1995.73.6.2563 . PMID 7666162 . - Перейти
↑ Moran, DW & Schwartz, AB (1999). «Моторное корковое представление скорости и направления при достижении».
J. Neurophysiol
.
82
(5): 2676–2692. DOI : 10,1152 / jn.1999.82.5.2676 . PMID 10561437 . - Kakei, С., Хоффман, Д. и Стрик P (1999). «Представления мышц и движений в первичной моторной коре». Наука
.
285
(5436): 2136–2139. CiteSeerX 10.1.1.137.8610 . DOI : 10.1126 / science.285.5436.2136 . PMID 10497133 . CS1 maint: несколько имен: список авторов ( ссылка ) - Перейти
↑ Todorov, E (2000). «Прямой корковый контроль мышечной активации при произвольных движениях рук: модель».
Nat Neurosci
.
3
(4): 391–398. DOI : 10.1038 / 73964 . PMID 10725930 . S2CID 13996279 . - ^ a b Haiss, F. & Schwarz, C (2005). «Пространственная сегрегация различных режимов управления движением в представлении усов первичной моторной коры крыс» . J. Neurosci
.
25
(6): 1579–1587. DOI : 10.1523 / JNEUROSCI.3760-04.2005 . PMC 6726007 . PMID 15703412 . - Ramanathan Д., Conner, JM и Tuszynski, MH (2006). «Форма моторной корковой пластичности, которая коррелирует с восстановлением функций после травмы головного мозга» . Proc. Natl. Акад. Sci. США
.
103
(30): 11370–11375. Bibcode : 2006PNAS..10311370R . DOI : 10.1073 / pnas.0601065103 . PMC 1544093 . PMID 16837575 . CS1 maint: несколько имен: список авторов ( ссылка ) - Брехта, М., Шнайдер, М., Sakmann, Б. и Margrie, TW (2004). «Движения усов, вызванные стимуляцией одиночных пирамидных клеток в моторной коре головного мозга крысы». Природа
.
427
(6976): 704–710. Bibcode : 2004Natur.427..704B . DOI : 10,1038 / природа02266 . PMID 14973477 . S2CID 1105868 . CS1 maint: несколько имен: список авторов ( ссылка ) - Перейти
↑ Cramer, NP & Keller, A (2006). «Кортикальный контроль центрального генератора паттернов взбалтывания» .
J. Neurophysiol
.
96
(1): 209–217. DOI : 10,1152 / jn.00071.2006 . PMC 1764853 . PMID 16641387 .
Дальнейшее чтение [ править ]
- Принципы неврологии (2000), 4-е изд., Kandel et al.
- Дебаэр, Ф., Вендерот, Н., Сунаерт, С., Ван-Хек, П., Суиннен, С.П. (июль 2003 г.). «Внутренняя и внешняя генерация движений: дифференциальные нервные пути, участвующие в бимануальной координации, выполняемые при наличии или отсутствии усиленной визуальной обратной связи». NeuroImage
.
19
(3): 764–76. DOI : 10.1016 / s1053-8119 (03) 00148-4 . PMID 12880805 .CS1 maint: несколько имен: список авторов ( ссылка ) - Воробьев; и другие. (1998). «Частичное разделение мезиальной области человека 6: цитоархитектонические свидетельства для трех отдельных областей». Eur J Neurosci
.
10
(6): 2199–203. DOI : 10,1046 / j.1460-9568.1998.00236.x . PMID 9753106 .