Подкорковые структуры головного мозга
Головной мозг является особо специализированной частью центральной нервной системы. У человека его масса составляет в среднем 1375 г. Именно здесь громадные скопления вставочных нейронов хранят полученный на протяжении жизни опыт действий. Головной мозг представлен 5-ю отделами. Три из них — продолговатый мозг, мост и средний мозг — объединяются под названием ствол (или — стволовая часть) головного мозга.
Стволовая часть принципиально отличается от двух других отделов мозга, так как снабжена черепными нервами, через которые ствол непосредственно контролирует область головы и часть шеи. Два других отдела — промежуточный и конечный мозг — не оказывают прямого влияния на структуры человеческого тела, они регулируют их функции, воздействуют на центры ствола и спинного мозга. Последние же, снабженные черепными и спинномозговыми нервами, передают через них обобщенные команды к исполнителям — мышцам и железам.
Помимо скоплений нейронов, имеющих прямое отношение к нервам, ствол мозга содержит и другие нервные центры, которые по характеру близки к центрам промежуточного и конечного мозга (ретикулярная формация, красные ядра, черная субстанция), что существенно отличает его от спинного мозга. Особое место занимает мозжечок, выполняющий важнейшие задачи по поддержанию степени напряжения мышц (тонуса), по координации их работы в выполнении движений, в поддержании равновесия при этом. В мозжечке громадное количество вставочных нейронов, которые он вмещает исключительно потому, что они находятся не только в его толще, но и в составе крайней складчатой поверхности, составляя кору мозжечка. Такой феномен проявляется, помимо него, только в коре конечного мозга.
Спереди и выше ствола находится промежуточный мозг и главными компонентами в виде зрительного бугра (таламуса) — важного промежуточного центра по ходу чувствительных путей к конечному мозгу, подбугорной области (гипоталамуса) — она содержит массу центров, важных для регуляции обмена веществ в организме, его поведения и тесно связана с функционированием гипофиза, с которым соединена ножкой. Позади зрительного бугра располагается эпифиз (шишковидное тело) — железа внутренней секреции, включенная в регуляцию пигментного обмена в коже и полового созревания.
Наибольшую часть массы головного мозга составляет конечный мозг, обычно описываемый как два полушария большого мозга, соединенные мозолистым телом. Его поверхность резко складчата из-за массы борозд (латеральная, центральная и др.), разделяющих извилины. Многие из них имеют постоянный характер, что позволяет различать участки коры.
Полушария разделяют на 4 основные доли. Лобная доля в значительной мере связана с определением личностных качеств человека, а ее задней части подчинены все двигательные центры ствола и спинного мозга. Поэтому при ее поражении появляются параличи мышц. В теменной доле, в основном, формируются ощущения тепла, холода, прикосновения, положения частей тела в пространстве. Затылочная доля содержит зрительные центры, височная — слуховые и обонятельные.
В глубине полушарий нейроны концентрируются в виде узлов (подкорки). Они вместе с другими центрами и мозжечком обеспечивают координацию работы мышц при выполнении двигательных программ разной сложности. Головной мозг окружен сложной системой оболочек. Мягкая оболочка сращена с его веществом и содержит в себе питающие мозг сосуды, ветви которых проникают в толщу мозга. Между ней и более поверхностной паутинной оболочкой, очень тонкой и бессосудистой, находится подпаутинное пространство со спинномозговой жидкостью. Ее большая часть продуцируется в полостях мозга (желудочков) и через отверстия между продолговатым мозгом и мозжечком выходит в это пространство, образуя вокруг мозга защитную гидравлическую подушку. Самая наружная твердая оболочка соединяется с костями черепа.
ПОДКОРКОВЫЕ СТРУКТУРЫ МОЗГА — отделы головного мозга, расположенные между корой больших полушарий и продолговатым мозгом. Оказывают активирующее влияние на кору, участвуют в формировании всех поведенческих реакций человека и животных, в поддержании мышечного тонуса и др.
К подкорковым образованиям относят структуры, находящиеся между корой головного мозга и продолговатым мозгом: таламус , гипоталамус, базальные ядра, комплекс образований, объединяемых в лимбическую систему мозга, а также ретикулярную формацию ствола мозга и таламуса. Любое афферентное возбуждение, возникшее при раздражении рецепторов на периферии, на уровне ствола мозга трансформируется в два потока возбуждений. Один поток по специфическим путям достигает специфической для данного раздражения проекционной области коры; другой — по коллатералям попадает в ретикулярную формацию, откуда в виде восходящего потока возбуждений направляется к коре больших полушарий, активируя ее. Ретикулярная формация имеет тесные функциональные и анатомические связи с гипоталамусом, таламусом, продолговатым мозгом, лимбической системой, мозжечком, поэтому многие виды деятельности организма (дыхание, пищевая и болевая реакции, двигательные акты и др.) осуществляются при ее обязательном участии.
Афферентные потоки возбуждений от периферических рецепторов на пути к коре больших полушарий имеют многочисленные синаптические переключения в таламусе. От латеральной группы ядер таламуса (специфические ядра) возбуждения направляются по двум путям: к подкорковым ганглиям и к специфическим проекционным зонам коры мозга. Медиальная группа ядер таламуса (неспецифические ядра) служит местом переключения восходящих активирующих влияний, которые направляются от стволовой ретикулярной формации в кору мозга. Тесные функциональные взаимосвязи между специфическими и неспецифическими ядрами таламуса обеспечивают первичный анализ и синтез всех афферентных возбуждений, поступающих в головной мозг. У животных, находящихся на низких ступенях филогенетического развития, таламус и лимбические образования играют роль высшего центра интеграции поведения, обеспечивая все необходимые двигательные акты животного, направленные на сохранение его жизни. У высших животных и человека высшим центром интеграции является кора больших полушарий.
К лимбической системе относят комплекс структур головного мозга, который играет ведущую роль в формировании основных врожденных реакций человека и животных: пищевых, половых и оборонительных. Он включает в себя поясничную извилину, гиппокамп, грушевидную извилину, обонятельный бугорок, миндалевидный комплекс и область перегородки. Центральное место среди образований лимбической системы отводится гиппокампу. Анатомически установлен гиппокампальный круг (гиппокамп -свод мозга- мамиллярные тела -передние ядра таламуса -поясная извилина -гиппокамп), который вместе с гипоталамусом играет ведущую роль в формировании эмоций. Регуляторные влияния лимбической системы широко распространяются на вегетативные функции (поддержание постоянства внутренней среды организма, регуляция кровяного давления, дыхания, тонуса сосудов, моторики желудочно – кишечного тракта, половых функций).
Кора больших полушарий оказывает постоянные нисходящие (тормозные и облегчающие) влияния на подкорковые структуры. Существуют различные формы циклического взаимодействия между корой и подкорковыми структурами, выражающиеся в циркуляции возбуждений между ними. Наиболее выраженная замкнутая циклическая связь существует между таламусом и соматосенсорной областью коры мозга, составляющими в функциональном отношении единое целое. Корково-подкорковая циркуляция возбуждений может служить основой для формирования условно — рефлекторной деятельности организма.
Таламус (thalamus, зрительный бугор) — структура, в которой происходит обработка и интеграция практически всех сигналов, идущих в кору большого мозга от спинного, среднего мозга, мозжечка, базальных ганглиев головного мозга.
Морфофункциональная организация. В ядрах таламуса происходит переключение информации, поступающей от экстеро-, проприорецепторов и интероцепторов и начинаются таламокортикальные пути.
Учитывая, что коленчатые тела таламуса являются подкорковыми центрами зрения и слуха, а узел уздечки и переднее зрительное ядро участвуют в анализе обонятельных сигналов, можно утверждать, что зрительный бугор в целом является подкорковой «станцией» для всех видов чувствительности. Здесь раздражения внешней и внутренней среды интегрируются, после чего поступают в кору большого мозга.
Зрительный бугор является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом (подтверждением тому служит наличие в таламусе около 120 разнофункциональных ядер). Ядра образуют своеобразные комплексы, которые можно разделить по признаку проекции в кору на 3 группы: передняя проецирует аксоны своих нейронов в поясную извилину коры большого мозга; медиальная — в лобную долю коры; латеральная — в теменную, височную, затылочную доли коры. По проекциям определяется и функция ядер. Такое деление не абсолютно, так как одна часть волокон от ядер таламуса идет в строго ограниченные корковые образования, другая — в разные области коры большого мозга.
Ядра таламуса функционально по характеру входящих и выходящих из них путей делятся на специфические, неспецифические и ассоциативные.
К специфическим ядрам относятся переднее вентральное, медиальное, вентролатеральиое, постлатеральное, постмедиальное, латеральное и медиальное коленчатые тела. Последние относятся к подкорковым центрам зрения и слуха соответственно.
Основной функциональной единицей специфических таламических ядер являются «релейные» нейроны, у которых мало дендритов и длинный аксон; их функция заключается в переключении информации, идущей в кору большого мозга от кожных, мышечных и других рецепторов.
От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры большого мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора большого мозга, имеют соматотопическую локализацию. Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же конвергируют сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Латеральное коленчатое тело имеет прямые эфферентные связи с затылочной долей коры большого мозга и афферентные связи с сетчаткой глаза и с передними буграми четверохолмий. Нейроны латеральных коленчатых тел по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
В медиальное коленчатое тело (МТК) поступают афферентные импульсы из латеральной петли и от нижних бугров четверохолмий. Эфферетные пути от медиальных коленчатых тел идут в височную зону коры большого мозга, достигая там первичной слуховой области коры. МКТ имеет четкую тонотопичность. Следовательно, уже на уровне таламуса обеспечивается пространственное распределение чувствительности всех сенсорных систем организма, в том числе сенсорных посылок от интерорецепторов сосудов, органов брюшной, грудной полостей.
Ассоциативные ядра таламуса представлены передним медиодорсальным, латеральным дорсальным ядрами и подушкой. Переднее ядро связано с лимбической корой (поясной извилиной), медиодорсальное — с лобной долей коры, латеральное дорсальное — с теменной, подушка — с ассоциативными зонами теменной и височной долями коры большого мозга.
Основными клеточными структурами этих ядер являются мультиполярные, биполярные трехотростчатые нейроны, т. е. нейроны, способные выполнять полисенсорные функции. Ряд нейронов изменяет активность только при одновременном комплексном раздражении. На полисенсорных нейронах происходит конвергенция возбуждений разных модальностей, формируется интегрированный сигнал, который затем передается в ассоциативную кору мозга. Нейроны подушки связаны главным образом с ассоциативными зонами теменной и височной долей коры большого мозга, нейроны латерального ядра — с теменной, нейроны медиального ядра — с лобной долей коры большого мозга.
Неспецифические ядра таламуса представлены срединным центром, парацентральным ядром, центральным медиальным и латеральным, субмедиальным, вентральным передним, парафасцикулярным комплексами, ретикулярным ядром, перивентрикулярной и центральной серой массой. Нейроны этих ядер образуют свои связи по ретикулярному типу. Их аксоны поднимаются в кору большого мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из РФ ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса.
Возбуждение неспецифических ядер вызывает генерацию в коре специфической веретенообразной электрической активности, свидетельствующей о развитии сонного состояния. Нарушение функции неспецифических ядер затрудняет появление веретенообразной активности, т. е. развитие сонного состояния.
Проекционными называют те волокна, которые связывают полушария головного мозга с нижележащими отделами мозга – стволом и спинным мозгом. В составе проекционных волокон проходят проводящие пути, несущие афферентную (чувствительную) и эфферентную (двигательную) информацию.
Итак, основные отделы, борозды и извилины головного мозга представлены на рис. 5.6.
Рис. 5. Головной мозг, левое полушарие (вид сбоку):
1 – прецентральная извилина; 2 – прецентральная борозда; 3 – верхняя лобная извилина; 4 – центральная борозда; 5 – средняя лобная извилина;
б – нижняя лобная извилина; 7 – восходящая ветвь латеральной борозды;
8 – горизонтальная ветвь латеральной борозды; 9 – задняя ветвь латеральной борозды; 10 – верхняя височная извилина; 11 – средняя височная извилина;
12 – нижняя височная извилина; 13 – теменная долька; 14 – пост-центральная борозда; 15 – постцентральная извилина; 16 – надкраевая извилина;
17 – угловая извилина; 18 – затылочная доля; 19 – мозжечок; 20 – горизонтальная щель мозжечка; 21 – продолговатый мозг
Т.В. Мельникова
Нижний Новгород, Россия
Изучение роли подкорковых структур и корково-подкоркового взаимодействия в организации высших психических функций (ВПФ) остается актуальной проблемой.
Входящие в состав активационного “энергетического” блока подкорковые образования имеют в то же время свои специфические клинические характеристики, и можно полагать, что они дифференцированно участвуют в реализации когнитивных функций.
Вместе с этим нейропсихологические исследования указывают на неоднородность активирующих систем, проявляющуюся снижением моторной, речевой, интеллектуальной активности при поражении левого полушария, особенностями эмоционально-личностной сферы при поражении правого.
Нарушения когнитивных функций при подкорковых поражениях зависят от распространенности патологического процесса и его этиологии (сосудистые или опухолевые заболевания мозга).
Данные о специфическом участии подкорковых ядер в психической деятельности, формировании межполушарной асимметри на подкорковом уровне получены при изучении ВПФ у больных паркинсонсонизмом до и после стереотаксических деструкций (Корсакова, Московичюте 1985).
Роль подкорковых структур в формировании когнитивных нарушений анализируется при прогрессирующей деменции (Корсакова и др., 1991), при болезни Паркинсона (Захаров и др., 1999), артери-венозных глубинных структурах мозга (Буклина, 1999).
Преобладание дисрегуляторных расстройств (при лакунарных инсультах зрительного бугра) выявлено при сопоставлении нарушений высших психических функций и МРТ (Яхно и др., 2002).
Зрительный бугор считается стратегически важной областью для реализации когнитивных функций, поэтому изучение нейропсихологического дефицита при поражениях различных его отделов представляет интерес для исследователей.
Нейропсихологическим исследованием при ограниченных таламических инсультах в отечественной и зарубежной литературе посвящено небольшое число публикаций. Во всех работах прослеживаются тенденции связывать возникновение когнитивных нарушений с поражением определенных сосудистых бассейнов и ядер зрительного бугра (Калашникова и др., 1998, Яхно и др., 2002).
В остром периоде, а также при двусторонних парамедианных поражениях отмечают снижение уровня бодрствования, сопор, нарушение памяти типа Корсаковского синдрома, галлюцинаторный синдром. Клинические проявления, наблюдаемые в подростковом периоде, рассматриваются как следствие нейродинамических нарушений.
При лакунаторных поражениях зрительного бугра в левом полушарии отмечают нарушения речевой функции (плохую артикуляцию, замедленность речи, парафазии), интеллектуальные, зрительно-пространственные, мнестические нарушения.
Сложная функция зрительного бугра обусловлена наличием специфических и неспецифических, вторичных и третичных групп ядер, а также многочисленными связями: соматосенсорными и ретикулотамическими путями, связями с орбитальной и префронтальной, теменной и затылочной корой, поясной извилиной, маммилярными телами. Типичный (задне-латеральный) таламический синдром Джерина-Русси, возникающий в результате закупорки таламо-коленчатой артерии, характеризуется преходящими контрлатеральным гемипарезом, контрлатеральной гемианестезией для тактильной и особенно глубокой чувствительности, гемигипестезией болевой и температурной чувствительности, спонтанной болью в вовлеченных областях, гемиатаксией, астереогнозом, контрлатеральным хореоатетозом.
Передне-боковой таламический синдром связан с закупоркой таламо-перфорирующей артерии и включает интенционный тремор, хореоатетоз, таламическую кисть.
Сопоставление результатов использования высокочувствительного нейропсихологического метода и данных КТ при лакунарных инсультах зрительного бугра позволяет оценить основные составляющие формирования нейропсихологического синдрома при этой патологии и возможность латерализации функций на этом уровне.
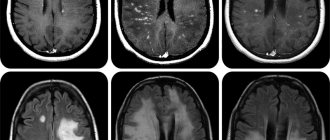
Лакунарные таламические инсульты по данным компьютерной томографии выявлены у 7 больных (5 женщин и 2 мужчин). Среднии возраст обследованных – 57 лет.
В трех случаях при компьютерной томографии головного мозга имелись указания на локализацию процесса в подушке зрительного бугра, в боковых и средних отделах, в 4 случаях локализация очага в пределах зрительного бугра точно не была определена. Правополушарная локализация была установлена у 5 больных, левополушарная у 2.
Обследование проводилось через 1-3 недели после острого инсульта. Ни в одном случае не было отмечено грубых нарушений памяти, дезориентации в месте и времени, галлюцинаторного синдрома. Только у одного больного был выявлен синдром Дежерина-Русси, с явлениями гемигипестезии, атаксии, жгучими болями в контрлатеральной половине тела. У остальных – очаговая симптоматика была легко выражена и проявлялась легкими преходящими гемипарезами, непостоянными нарушениями чувствительности, атаксией.
асимметрия реакции полушарий – предыдущая | следующая – зрительный бугор
А. Р. Лурия и психология XXI века. Содержание
Строение подкорковой области головного мозга. Промежуточный мозг
Стриопаллидарная система
В толще белого вещества полушарий мозга располагаются скопления серого вещества, называемые подкорковыми ядрами (базальные ядра). К ним относятся хвостатое ядро, чечевицеобразное ядро, ограда и миндалевидное тело (рис. 6). Чечевицеобразное ядро, находящееся снаружи от хвостатого ядра, делится на три части. В нем различают скорлупу и два бледных шара.
Рис. 6. Подкорковые ядра:
1 – хвостатое ядро; 2 – чечевицеобразное ядро; 3 – зрительный бугор.
А – горизонтальный разрез: а – ограда; б – скорлупа; в и г – бледный шар;
В-фронтальный разрез: а – бледный шар; б – скорлупа
В функциональном отношении хвостатое ядро и скорлупа объединяются в полосатое тело (стриатум), а бледные шары вместе с черной субстанцией и красными ядрами, расположенными в ножках мозга, – в бледное тело (паллидум).
Вместе они представляют очень важное в функциональном отношении образование – стриоппаллидарную систему. По морфологическим особенностям и филогенетическому происхождению (появление их на определенной ступени эволюционного развития) бледное тело является более древним, чем полосатое тело, образованием.
Стриопаллидарная система является важной составной частью двигательной системы. Она входит в состав так называемой пирамидной системы. В двигательной зоне коры головного мозга начинается двигательный – пирамидный – путь, по которому следует приказ выполнить то или иное движение.
Экстрапирамидная система, важной составной частью которой является стриопаллидум, включаясь в двигательную пирамидную систему, принимает подсобное участие в обеспечении произвольных движений.
В то время, когда кора головного мозга еще не была развита, Стриопаллидарная система была главным двигательным центром, определявшим поведение животного. За счет стриопаллидарного двигательного аппарата осуществлялись диффузные, массовые, движения тела, обеспечивающие передвижение, плавание и т.п.
С развитием коры головного мозга Стриопаллидарная система перешла в подчиненное состояние. Главным двигательным центром стала кора головного мозга.
Стриопаллидарная система стала обеспечивать фон, готовность к совершению движения; на этом фоне осуществляются контролируемые корой головного мозга быстрые, точные, строго дифференцированные движения.
Для совершения движения необходимо, чтобы одни мышцы сократились, а другие расслабились, иначе говоря, нужно точное и согласованное перераспределение мышечного тонуса.
Такое перераспределение тонуса мышц как раз и осуществляется стриопаллидарной системой. Эта система обеспечивает наиболее экономное потребление мышечной энергии в процессе выполнения движения. Совершенствование движения в процессе обучения их выполнению (например, отработка до предела отточенного бега пальцев музыканта, взмаха руки косаря, точных движений водителя автомобиля) приводит к постепенной экономизации и автоматизации.
Такая возможность обеспечивается стриопаллидарной системой.
Выше было отмечено, что в филогенетическом отношении полосатое тело – образование более молодое, чем бледное тело. Примером паллидарных организмов являются рыбы.
Они передвигаются в воде с помощью бросковых мощных движений туловища, не «заботясь» об экономии мышечной энергии. Эти движения имеют относительно точный и мощный характер. Однако они расточительны энергетически. У птиц полосатое тело уже хорошо выражено, что помогает им более расчетливо регулировать качество, точность и количество движений. Таким образом, бледное тело тормозит и регулирует деятельность паллидарный системы (т. к. филогенетически более молодые образования контролируют и тормозят более древние).
Двигательные акты новорожденного носят паллидарный характер: они некоординированны, бросковы и часто излишни. С возрастом, по мере созревания стриатума, движения ребенка становятся более экономичными, скупыми, автоматизированными.
Стриопаллидарная система имеет связи с корой головного мозга, корковой двигательной системой (пирамидной) и мышцами, образованиями экстрапирамидной системы, со спинным мозгом и зрительным бугром.
Другие базальные ядра (ограда и миндалевидное тело) расположены кнаружи от чечевицеобразного ядра. Миндалевидное тело входит в другую функциональную систему – лимбико-ретикулярный комплекс.
Зрительный бугор
Из промежуточного мозгового пузыря развиваются зрительный бугор и Подбугорная область (гипоталамус), из полости промежуточного мозгового пузыря – III желудочек.
Зрительный бугор,
или
таламус,
расположен по сторонам III желудочка и состоит из мощного скопления серого вещества.
Зрительный бугор делят на собственно зрительный бугор, надбугорную (надталамическую область, или эпиламус) и забугорную (заталамическая область, или метаталус). Основную массу серого бугра составляет таламус (см. рис. 7).
Рис. 7. Топография таламуса
1 – таламус; 2 – тело хвостатого ядра; 3 – тело бокового желудочка; 4 – мозолистое тело; 5 – продолговатый мозг.
В нем выделяют выпячивание подушку, кзади от которой имеются два возвышения – наружное и внутреннее коленчатые тела (они входят в забугорную область).
В таламусе различают несколько ядерных групп.
Надбугорная область, или эпиталамус состоит из шишковидной железы и задней спайки мозга.
Забугорная область, или метаталамус, включает в себя коленчатые тела, являющиеся возвышением таламуса. Они лежат кнаружи и книзу от подушки таламуса.
Подбугорная область, или гипоталамус, лежит книзу от таламуса, имеет ряд ядер, лежащих в стенках III желудочка.
Зрительный бугор является важным этапом на пути проведения всех видов чувствительности. К нему подходят и в нем сосредоточиваются чувствительные пути – осязание, болевое, температурное чувство, зрительные тракты, слуховые пути, обонятельные пути и волокна от экстрапирамидной системы. От нейронов зрительного бугра начинается следующий этап передачи чувствительных импульсов – в кору головного мозга.
На определенном этапе эволюции нервной системы таламус был центром чувствительности, подобно тому, как Стриопаллидарная система – механизмом движений. По мере появления и развития коры головного мозга основная роль в функции чувствительной сферы перешла коре головного мозга, а зрительный бугор остался лишь передаточной станцией чувствительных импульсов от периферии к коре мозга.