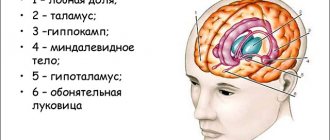
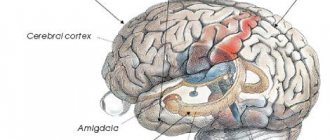
Структуры, связанные с лимбической системой, расположены во внутренней части височной доле мозга: gyrus hippocampalis, рострально переходящие в uncus и включающие в себя амигдал и гиппокамп, участвуют в регуляции функций вегетативной нервной системы, аффективной сферы. Этим участкам мозга также приписывают ответственность за побуждения и мотивацию, память и возможность обучения.
Лимбическая система мозга, как известно, связана с эмоциональной сферой человека, ее повреждение при шизофрении вполне допустимо, учитывая колебания настроения, маниакальные состояния, депрессию и ажиатацию, наблюдаемые при этом заболевании.
Возможно, при шизофрении страдают не все компоненты лимбической системы в равной степени, однако в строении нейронов, входящих в эту систему, могут иметь место более тонкие морфологические изменения. Некоторые, но все же отличные из этих изменений в ряде случаев были обнаружены у больных, страдающих биполярным аффективным расстройством.
Результаты многих исследований при шизофрении показывают уменьшение плотности нейронов в области гипоталамуса, комплекса миндалина — гиппокамп и парагиппокампальной извилины.
Размеры парагиппокампальной извилины и энторинального кортекса, согласно K. Prasad et al. (2004), обнаруживают определенную корреляцию с выраженностью бреда и психотической симптоматики при шизофрении. По мнению исследователей, эти структуры мозга играют важную роль в регуляции процессов, связанных с памятью.
Обнаружено сокращение размеров и изменение формы нейронов в гиппокампе, гиппокампальной извилине и энторинальной области коры мозга (Bogerts B., 1993).
Гиппокамп
В исследовании C. McDonald et al. (2006) было выявлено уменьшение объема правого и левого гиппокампа, 2,47 мл и 2,50 мл, против 2,54 мл и 2,61 мл у здоровых лиц, что соответствует примерно уменьшению объема этой структуры мозга примерно на 6%.
Некоторые авторы отмечают, что уменьшение объема гиппокампа, как части лимбической системы, заметно уже после первого психотического эпизода, однако, по мнению других исследователей, эти изменения фиксируются и до манифестации шизофрении и прогрессируют после ее начала. Отметим, что у родственников больных шизофренией также можно выявить уменьшение объема гиппокампа и амигдала.
При шизофрении нарушено функционирование амигдала, этот факт обнаруживается при исследовании больных шизофренией с помощью метода вызванных потенциалов (Р300).
В гиппокампе больных шизофренией при уменьшении объема нейронов, «разряженности» их взаиморасположения обнаружено увеличение числа патологически измененных миелинизированных аксонов с истонченными миелиновыми оболочками, набухшими периоксанальным глиальным отростком и сморщенным аксоном. Пропорция этих волокон в общем числе миелинизированных аксонов и их численная плотность при шизофрении в 2 раза больше, чем в контрольной группе. В то же время известно, что патологические и репаративные изменения аксонов зависят от реакции, окружающих их клеток микроглии (Коломеец Н.С., 2007).
Своеобразные нарушения памяти, в первую очередь рабочей, при шизофрении, сказывающиеся на трудностях обучения лиц, страдающих этой болезнью могут указывать на вовлечение гиппокампа в патологический процесс.
По данным Shenton et al. (2001), объемы медиальных отделов височных долей, обычно включающих в себя гиппокамп и миндалевидное тело, значительно сокращены в своих размерах. Эти изменения были отмечены у 70% больных шизофренией.
Лимбическая система ( продолжение)
По мнению James, эмоции связаны с активностью сенсорного неокортекса, как места интеграции ощущений тела воспроизведенных в эмоциональных состояниях. Тот факт, точнее концепция о том, что определенные структуры мозга могут формировать эмоциональное состояния ( эмоциональные системы мозга) было несколько неожиданным для неврологов и по сути явилось своего рода стартовой площадкой такой дисциплины, как биологическая психиатрия и «поведенческая неврология» — ветви неврологии , ксающейся того , как неврологические расстройства связаны с изменениями поведения.
В 1937 году Papez описал предполгаемый церебральный механизм эмоций . Ключевым компонентом его сегодня хорошо известного круга были гиппокамп , поясная извилина, маммилярные тела и передний таламус. Для Papez цингулярный кортекс являлся рецептивной областью для переживаемых эмоций, таким же образом , как визуальная область мозга являлась рецептивной для визуальной информации. Гипоталамус являлся основой для выражения ( экспрессии) эмоций, а для их переживания необходима была кора мозга , «поток чувств» зависил от силы связей между кортексом и гипоталамусом. В заключении Papez обобщил , сегодня широко известной цитатой , что «гипоталамус , передние таламические ядра , поясная извилина , гиппокамп и их взаимосвязи образуют гармонический механизм , который может представлять собой сложую в функциональном плане центральную эмоцию и также учавствующую в эмоциональном переживании.
Ключевыми структурами лимбической системы оригинально обрисованной в общих чертах MacLean , но детально разработанной другими авторами , были миндалина и гиппокамп — своего рода нейронные аггрегаты функционирующие в комплексе с непосредственно связанными с ними структурами , такими как орбитальная часть фронтальной коры и так называемый вентральный стриатум. — часть экстрапирамидной системы мозга , которая имеет отношение к двигательной составляющей эмоции ( «моторное выражение или экспрессия эмоции»). У человека в связи с миграцией височной доли к задне — нижним отделам мозга и большого размера мозолистого тела архикортекс ( эволюционно старый кортекс ) , находится как бы в свернутом виде и закрытом в медиальной части височной доли — в гиппокампе.
Поясная извилина окружает мозолистое тело , формируя С — образную группу , соединенную в задней части с парагиппокампальной извилиной и экстенсивно контактирующей с неокортикальными структурами. Лобная доля мозга включает в себя большое количество демаркацинных частей ( субрегионов) , но наиболее активны и известны орбитальная , медиальная и дорсолатеральная области фронтальной доли. Орбитофронтальный кортекс расположен выше черепной ямки и имеет интимные связи с передним островком , миндалиной , вентральным стриатумом и трактами сенсорных проекций. Вентромедиальный фронтальный кортекс представляет собой часть поясной извилины . Островок — сравнительно большая лимбическая структура , которая по контрасту с большинством компонентов лимбической системы остается невидимой со стороны медиальной поверхности мозга и лежит латерально и заключена ( закрыта) ниже изгибов неокортекса.
Многие исследователи критично выссказываются в отношении термина «лимбическая система» , а некоторые анатомы сегодня предпочитают пользоваться термином «лимбическая доля» . после оригинального обозначения Broca тем самым как бы разрушая концепцию «лимбической системы». С развитием таких современных техник нейроанатомии , как использование пероксидазы хрена ( HRP) , гистофлюоресценции, ауторадиографии и модернизированный метод окрашивания серебром , предложенный еще на ранних этапах развития нейроанатомии , заметно вырос интерес к структурно — функциональным особенностям лимбической доли, особенно кортикально — субкортикальных взаимосвязей , в частности, в плане развития представлений о нейроанатомии эмоций и поведения.
Несмотря на то, что термин Broca в большей степени является топографическим , лимбическая доля может быть дифференцирована ( отделена ) от других кортикальных структур окрашиванием клеток и уязвимостью по отношению к некоторым психоневрологическим расстройствам. В одном из последних определений , в частности предложенных Heimer et.al. ( 2007) , лимбическая доля состоит из ольфакторного аллокортекса, гипокампального аллокортекса, переходных областей коры, которые осуществляют связи с большим изокортексом. Переходные области достаточно многочисленны и формируют большую часть лимбической доли и отедлены в понятиях «структура» одной и более границами ( трактами) от изокортекса. Кортикальные поля лимбической доли образуют поясную и парагиппокампальную извилины , хорошо связывая или соединяя каудальную орбитофронтальную , медиальную фронтальную , височно — полярную , антеровентральную часть островка и ретросплениальный кортекс, а также включают в себя латеробазально — кортикальный комплекс миндалины.
Расширенная оригинальная конпцеия лимбической системы была предложена Nauta , Domenesick (1982) , которые включили задний орбито — фронтальный кортекс и височный полюс , отдающий проекции к миндалине и гиппокампу , а также некоторым субкортикальным структурам , связанных между собой реципрокными путями. Они включают в себя аккумбенс ( прилежащие ядра) и такие структуры среднего мозга , как вентральная область покрышки ( VTA) и интепедункулярные ядра . Включения этих структур привели к образованию понятия «среднемозговая лимбическая область» «мезолимбической системы».
Nienwenhuys ( 1996) в свое время ввел термин «большая лимбическая система» , которая также включал в себя мост и медуллярную нейрональную группу , имеющую отношение к вегетативной и сомато — моторной интеграции. Он подчеркнул, что данная система состоит из массы тоникх и ультратонких волокон ( фибр) и включающей в себя нейрональные группы , имеющие отношение к моноаминовой и пептидной активности и содержит в себе места ( очаги) , с активностью которых могут быть ассоциированы некоторые паттерны поведения.
Важно понимать, что анатомически лимбическая доля представляет собой не изокортикальную структуру , не имеет шестислойную структуру , а состит из аллокортекса и пояса агранулярного или дизламинарного ( 4 поле отсутствует , плохо организовано или атипично) промежуточного кортекса. Другими словами, возможные определения лимбического кортекса связаны с теми областями коры , которые не являются изокортикальными ( шестислойными).
Входные пути к лимбической системе могут быть как интероцептивными ( висцеральными) , так и экстерорецептивными ( передающих информацию из окружающей среды). Формирование из разных структур предоставляют информацию о внутренней среде организма и включают в себя модулирующее влияние нейротрансмиттерных трактов , которые происходят из среднего и заднего мозга и обеспечивают драйв поведения и модуляцию эмоционального состояния с помощью дофамина, серотонина и норадреналина. Экстерорецептивные афферентные связи от всех сенсорных систем в конечном счете, комплексно интегрируют сенсорную информацию для кортекса гиппокампа и миндалины.
Миндалевидные ядра
В серии аутопсий головного мозга больных шизофренией не выявлено существенного увеличения объема миндалевидных ядр (Steven A. et al., 2002). Однако, данные полученные при помощи МРТ, в настоящее время относятся к достаточно грубым методам изображения, не говоря уже о КТ и по сравнению с аутопсией могут быть менее достоверными.
В сравнительном обзоре Wright et al (2000), изучавшим размеры 44 областей мозга при шизофрении, было отмечено, что миндалины левого и правого полушарий сокращены в своем объеме на 10% и это заметно в большей степени, чем сокращение обема остальных участков мозга. Потеря серого вещества миндалевидного тела и изменение формы последнего так же были доказана с помощью тех исследований, которые обращали внимание на показатели желудочко-мозгового индекса. В то же время многие авторы отмечают методологические трудности исследования миндалевидного тела лимбической системы больных шизофренией.
Вернуться к Содержанию
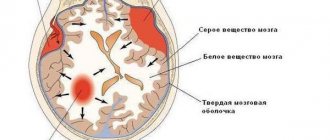
Белое вещество
Оно представлено в виде многочисленных волокон. Их разделяют на три группы:
- Проекционные. Эта категория представлена пучками эфферентных и афферентных волокон. Посредством них существуют связи между проекционными центрами и базальными, стволовыми и спинномозговыми ядрами.
- Ассоциативные. Эти волокна обеспечивают соединение корковых участков в границах одного полушария. Они разделены на короткие и длинные.
- Комиссуральные. Эти элементы связывают корковые зоны противоположных полушарий. Комиссуральными образованиями считаются: мозолистое тело, задняя и передняя спайка и спайка свода.
Поверхность плащевого элемента
В каждом сегменте эта часть мозга разделена на доли посредством глубоких борозд и щелей. Первичные относят к постоянным образованиям органа. Формируются они на эмбриональном этапе (на пятом месяце). К наиболее крупным щелям относят продольную (разделяет сегменты) и поперечную (отделяет от затылочных долей мозжечок).
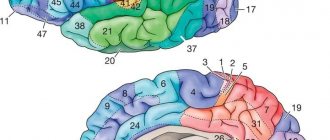
Вторичными и в особенности – третичными образованиями определяется индивидуальный рельеф сегментов (он просматривается на фото). Мозг человека развивается не только во внутриутробный период. К примеру, вторичные и третичные борозды формируются до 7-8 лет после рождения. Рельеф, который имеет конечный мозг, местонахождение постоянных образований и крупных извилин у большинства людей схожи. В каждом сегменте выделяют по шесть долей: лимбическую, островковую, височную, затылочную, теменную и лобную.
Конечный мозг в этой области включает в себя роландову (центральную) борозду. С ее помощью разделяются теменная и лобная доли. Также на поверхности присутствует сильвиева (боковая) борозда. Посредством нее теменная и лобная доли отделены от височной. В качестве передненижней границы затылочной области выступает условная линия.
Неокортекс
Новая кора – часть мозга, присущая высшим млекопитающим. Зачатки неокортекса также наблюдаются у низших животных, сосущих молоко, однако они не достигают высокого развития. У человека изокортекс – львиная часть общей коры головного мозга, имеющая толщину в среднем до 4 миллиметров. Площадь неокортекса достигает 220 тысяч кв. мм.
История возникновения
В данный момент неокортекс – высшая ступень эволюции человека. Первые проявления новой коры ученым удалось изучить у представителей рептилий. Последними животными, не имеющие новой коры в цепочке развития, оказались птицы. И лишь развитой нейронной системой обладает человек.
Эволюция — сложный и длинный процесс. Каждый вид существ проходит суровый эволюционный процесс. Если вид животного не смог адаптироваться под изменчивую внешнюю среду — вид терял свое существование. Почему же человек смог адаптироваться и выжить по сей день ?
Находясь в благоприятных условиях проживания (теплый климат и белковая еда), потомкам человека (до Неандертальцев) не оставалось ничего, как питаться и размножаться ( благодаря развитой лимбической системе). Из-за этого масса мозга, по меркам длительности эволюции, набрала критическую массу за небольшой период времени (несколько миллионов лет). Кстати, масса мозга в те времена была на 20% больше, чем у современного человека.
Однако, всему хорошему рано или поздно приходит конец. Со сменой климата, потомкам нужно было менять место жительство, а с ним и начинать искать еду. Имея огромный мозг, потомки начали применять его для поиска пищи, а далее и для социального вовлечения, т.к. выяснилось, что объединяясь в группы по определенным критериям поведения — выживать было легче. К примеру, в группе, где каждый делился пищей с другими членами группы имела больше шансов на выживание (Кто-то хорошо собирал ягоды, а кто-то охотился и тд).
С этого момента началась отдельная эволюция по мозгу, отдельная от эволюции всего тела. С тех времен внешний вид человека не сильно поменялся, но состав мозгов отличается кардинально.
Из чего состоит
Новая кора больших полушарий – это скопление нервных клеток, образующих комплексное серое вещество. Анатомически разделяют 4 типа коры, в зависимости от её локализации – теменная, затылочная, лобная, височная. Гистологически же кора состоит из шести шаров клеток:
- Молекулярный шар;
- наружный зернистый;
- пирамидные нейроны;
- внутренний зернистый;
- ганглионарный слой;
- мульиформные клетки.
Какие функции выполняет
Новая кора головного мозга человека классифицируется по трем функциональным зонам:
- Сенсорная. Эта зона отвечает за высшую обработку полученных раздражителей из внешней среды. Так, лед становится холодным тогда, когда информация о температуре поступает в теменную область – на пальце же холода нет, а есть только электрический импульс.
- Ассоциативная зона. Эта область коры отвечает за информационную связь между моторной корой и чувствительной.
- Моторная зона. В этой части мозга формируются все сознательные движение. Кроме таких функций, новая кора обеспечивает высшую психическую деятельность: интеллект, речь, память и поведение.
Нервные центры коры больших полушарий головного мозга
Участки коры полушарий, имеющие характерную цитоархитектонику, и нервные связи, участвующие в выполнении определенных функций, являются нервными центрами. Поражение таких участков коры проявляется в утрате присущих им функций. Нервные центры плаща могут быть разделены на проекционные и ассоциативные.
Проекционные центры коры больших полушарий головного мозга — это участки, представляющие собой корковую часть анализатора, имеющие непосредственную морфофункциональную связь через посредство афферентных или эфферентных нервных путей с нейронами подкорковых центров.
Ассоциативные центры — это участки коры больших полушарий головного мозга человека, не имеющие непосредственной связи с подкорковыми образованиями, а связанные временной двусторонней связью с проекционными центрами. Ассоциативные центры играют первостепенную роль в осуществлении высшей нервной деятельности. В настоящее время достаточно точно выяснена динамическая локализация некоторых функций коры полушарий большого мозга. Участки коры полушарий большого мозга, не являющиеся проекционными или ассоциативными центрами, участвуют в выполнении межанализаторной интегративной деятельности головного мозга.
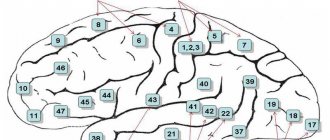
Корковые поля неравноценны в функциональном отношении и их можно разделить на первичные, вторичные и третичные.
Первичные поля представляют собой четко отграниченные участки, которые соответствуют центральным частям анализаторов. В эти поля проходит по специфическим проекционным афферентным путям основная масса сигналов от органов чувств. Первичные поля характеризуются сильным развитием внутренней зернистой пластинки. Первичные поля связаны с реле-ядрами таламуса и ядрами коленчатых тел. Они имеют экранную структуру и, как правило, жесткую соматотопическую проекцию, при которой отдельные участки периферии проецируются в соответствующие им участки коры. Повреждение первичных полей коры сопровождается нарушением непосредственного восприятия и тонкой дифференцировки раздражений.
Вторичные поля коры примыкают к первичным полям. Их можно рассматривать как периферические части корковых анализаторов. Эти поля связаны с ассоциативными ядрами таламуса. При поражении вторичных полей сохраняются элементарные ощущения, но нарушается способность к более сложным восприятиям. Вторичные поля не имеют четких границ, в них не выражена соматотопическая проекция.
Третичные поля коры отличаются наиболее тонкой нейронной структурой и преобладанием ассоциативных элементов. Эти поля связаны сзадними ядрами таламуса. В третичных полях осуществляются наиболее сложные взаимодействия анализаторов, лежащие в основе познавательного процесса (гнозия), формируются программы целенаправленных действий (праксия).
Кора обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций и имеет следующие морфофункциональные особенности:
- Многослойность расположения нейронов;
- Модульный принцип организации;
- Соматотопическая локализация рецептирующих систем;
- Экранность, т.е. распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора;
- Зависимость уровня активности от влияния подкорковых структур и ретикулярной формации;
- Наличие представительства всех функций нижележащих структур ЦНС;
- Цитоархитектоническое распределение на поля;
- Наличие в специфических проекционных сенсорных и моторной системах вторичных и третичных полей с ассоциативными функциями;
- Наличие специализированных ассоциативных областей;
- Динамическая локализация функций, выражающаяся в возможности компенсаций функций утраченных структур;
- Перекрытие в коре большого мозга зон соседних периферических рецептивных полей;
- Возможность длительного сохранения следов раздражения;
- Реципрокная функциональная взаимосвязь возбудительных и тормозных состояний;
- Способность к иррадиации (распространению) возбуждения и торможения;
- Наличие специфической электрической активности.
Важно Всд по ваготоническому типу