Функции стриопаллидарной системы, лимбической системы и коры полушарий большого мозга
№1.1. Функции бледного шара. Афферентные и эфферентные связи
Бледный шар (globus pallidus s. pallidum) имеет преимущественно крупные нейроны Гольджи I типа. Связи бледного шара с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом, соматосенсорной системой и др. свидетельствуют об его участии в организации простых и сложных форм поведения.
Раздражение бледного шара с помощью вживленных электродов вызывает сокращение мышц конечностей, активацию или торможение γ-мотонейронов спинного мозга. У больных с гиперкинезами раздражение разных отделов бледного шара (в зависимости от места и частоты раздражения) увеличивало или снижало гиперкинез.
Стимуляция бледного шара в отличие от стимуляции хвостатого ядра не вызывает торможения, а провоцирует ориентировочную реакцию, движения конечностей, пищевое поведение (обнюхивание, жевание, глотание и т.д.).
Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи. При повреждении бледного шара наблюдается миоклония — быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица.
В первые часы после повреждения бледного шара в остром опыте на животных резко снижалась двигательная активность, движения характеризовались дискоординацией, отмечалось наличие незавершенных движений, при сидении — поникшая поза. Начав движение, животное долго не могло остановиться. У человека с дисфункцией бледного шара затруднено начало движений, исчезают вспомогательные и реактивные движения при вставании, нарушаются содружественные движения рук при ходьбе, появляется симптом пропульсии: длительная подготовка к движению, затем быстрое движение и остановка. Такие циклы у больных повторяются многократно.
Большая часть афферентных сигналов, приходящих к базальным ганглиям, поступает в полосатое тело базальных ганглий. Эти сигналы исходят почти исключительно из трех источников: от всех областей коры больших полушарий, от внутрипластинчатых ядер таламуса и черной субстанции базальных ганглий. Эфферентные волокна от стриатума идут к бледному шару базальных ганглий и черной субстанции, от которой начинается дофаминергический путь к полосатому телу и пути, идущие к таламусу. От внутреннего отдела бледного шара берет начало самый важный из всех эфферентных трактов, заканчивающийся в таламусе и в крыше среднего мозга.
№1.2. Функции хвостатого ядра и скорлупы. Афферентные и эфферентные связи
Хвостатое ядро (nucleus caudatus) и скорлупа (putamen) являются эволюционно более поздними, чем бледный шар, образованиями и функционально оказывают на него тормозящее влияние.
Хвостатое ядро и скорлупа имеют сходное гистологическое строение. Их нейроны относятся ко II типу клеток Гольджи, т. е. имеют короткие дендриты, тонкий аксон; их размер до 20 мк. Этих нейронов в 20 раз больше, чем нейронов Гольджи I типа, имеющих разветвленную сеть дендритов и размер около 50 мк.
Функции любых образований головного мозга определяются прежде всего их связями, которых у базальных ядер достаточно много. Эти связи имеют четкую направленность и функциональную очерченность.
Хвостатое ядро и скорлупа получают нисходящие связи преимущественно от экстрапирамидной коры через подмозолистый пучок. Другие поля коры большого мозга также посылают большое количество аксонов к хвостатому ядру и скорлупе.
Основная часть аксонов хвостатого ядра и скорлупы идет к бледному шару, отсюда — к таламусу и только от него — к сенсорным полям. Следовательно, между этими образованиями имеется замкнутый круг связей. Хвостатое ядро и скорлупа имеют также функциональные связи со структурами, лежащими вне этого круга: с черной субстанцией, красным ядром, люисовым телом, ядрами преддверия, мозжечком, γ -клетками спинного мозга.
Обилие и характер связей хвостатого ядра и скорлупы свидетельствуют об их участии в интегративных процессах, организации и регуляции движений, регуляции работы вегетативных органов.
Раздражение поля 8 коры большого мозга вызывает возбуждение нейронов хвостатого ядра, а поля 6 — возбуждение нейронов хвостатого ядра и скорлупы. Одиночное раздражение сенсомоторной области коры большого мозга может вызывать возбуждение или торможение активности нейронов хвостатого ядра. Эти реакции возникают через 10—20 мс, что свидетельствует о прямых и опосредованных связях коры большого мозга с хвостатым ядром.
Медиальные ядра таламуса имеют прямые связи с хвостатым ядром, свидетельством чего служит реакция его нейронов, наступающая через 2—4 мс после раздражения таламуса.
Реакцию нейронов хвостатого ядра вызывают раздражения кожи, световые, звуковые стимулы. —
Во взаимодействиях хвостатого ядра и бледного шара превалируют тормозные влияния. Если раздражать хвостатое ядро, то большая часть нейронов бледного шара тормозится, а меньшая возбуждается. В случае повреждения хвостатого ядра у животного появляется двигательная гиперактивность.
Взаимодействие черного вещества и хвостатого ядра основано на прямых и обратных связях между ними. Установлено, что стимуляция хвостатого ядра усиливает активность нейронов черного вещества. Стимуляция черного вещества приводит к увеличению, а разрушение — к уменьшению количества дофамина в хвостатом ядре. Установлено, что дофамин синтезируется в клетках черного вещества, а затем со скоростью 0,8 мм/ч транспортируется к синапсам нейронов хвостатого ядра. В хвостатом ядре в 1 г нервной ткани накапливается до 10 мкг дофамина, что в 6 раз больше, чем в других отделах переднего мозга, бледном шаре, в 19 раз больше, чем в мозжечке. Благодаря дофамину проявляется растормаживающий механизм взаимодействия хвостатого ядра и бледного шара.При недостатке дофамина в хвостатом ядре (например, при дисфункции черного вещества) бледный шар растормаживается, активизирует спинно-стволовые системы, что приводит к двигательным нарушениям в виде ригидности мышц.
Кортико-стриарные связи топически локализованы. Так, передние области мозга связаны с головкой хвостатого ядра. Патология, возникающая в одной из взаимосвязанных областей кора — хвостатое ядро, функционально компенсируется сохранившейся структурой.Кортико-стриарные связи топически локализованы. Так, передние области мозга связаны с головкой хвостатого ядра. Патология, возникающая в одной из взаимосвязанных областей кора — хвостатое ядро, функционально компенсируется сохранившейся структурой.
Хвостатое ядро и бледный шар принимают участие в таких интегративных процессах, как условнорефлекторная деятельность, двигательная активность. Это выявляется при стимуляции хвостатого ядра, скорлупы и бледного шара, деструкции и при регистрации электрической активности.
Прямое раздражение некоторых зон хвостатого ядра вызывает поворот головы в сторону, противоположную раздражаемому полушарию, животное начинает двигаться по кругу, т. е. возникает так называемая циркуляторная реакция.
Раздражение других областей хвостатого ядра и скорлупы вызывает прекращение всех видов активности человека или животного: ориентировочной, эмоциональной, двигательной, пищевой. При этом в коре большого мозга наблюдается медленноволновая активность.
У человека стимуляция хвостатого ядра во время нейрохирургической операции нарушает речевой контакт с больным: если больной что-то говорил, то он замолкает, а после прекращения раздражения не помнит, что к нему обращались. В случаях травм головного мозга с раздражением головки хвостатого ядра у больных отмечается ретро-, антеро- или ретроантероградная амнезия.
У таких животных, как обезьяны, раздражения хвостатого ядра на разных этапах реализации условного рефлекса приводят к торможению выполнения данного рефлекса. Например, если у обезьяны через вживленные электроды раздражать хвостатое ядро перед подачей условного сигнала, то обезьяна не реагирует на сигнал, как будто не слышала его; раздражение ядра после того, как обезьяна на сигнал направляется к кормушке или уже начинает брать пищу из кормушки, приводит к остановке животного, после прекращения раздражения обезьяна, не завершив условной реакции, возвращается на место, т. е. она «забывает», что был раздражающий сигнал (ретроградная амнезия).
Раздражение хвостатого ядра может полностью предотвратить восприятие болевых, зрительных, слуховых и других видов стимуляции. Раздражение вентральной области хвостатого ядра снижает, а дорсальной — повышает слюноотделение.
При стимуляции хвостатого ядра удлиняются латентные периоды рефлексов, нарушается переделка условных рефлексов. Выработка условных рефлексов на фоне стимуляции хвостатого ядра становится невозможной. Видимо, это объясняется тем, что стимуляция хвостатого ядра вызывает торможение активности коры большого мозга.
Ряд подкорковых структур также получает тормозное влияние хвостатого ядра. Так, стимуляция хвостатых ядер вызывала веретенообразную активность в зрительном бугре, бледном шаре, субталамическом теле, черном веществе и др.
Таким образом, специфичным для раздражения хвостатого ядра является преимущественно торможение активности коры большого мозга, подкорковых образований, торможение безусловного и условнорефлекторного поведения.
В то же время при раздражении хвостатого ядра могут появляться некоторые виды изолированных движений. Видимо, хвостатое ядро имеет наряду с тормозящими и возбуждающие структуры.
Выключение хвостатого ядра сопровождается развитием гиперкинезов типа непроизвольных мимических реакций, тремора, атетоза, торсионного спазма, хореи (подергивания конечностей; туловища, как при некоординированном танце), двигательной гиперактивности в форме бесцельного перемещения с места на место.
В случае повреждения хвостатого ядра наблюдаются существенные расстройства высшей нервной деятельности, затруднение ориентации в пространстве, нарушение памяти, замедление роста организма. После двустороннего повреждения хвостатого ядра условные рефлексы исчезают на длительный срок, выработка новых рефлексов затрудняется, общее поведение отличается застойностью, инертностью, трудностью переключений. У обезьян после одностороннего повреждения хвостатого ядра условная реакция восстанавливалась через 30—50 дней, латентные периоды рефлексов удлинялись, появлялись межсигнальные реакции. Двустороннее повреждение приводило к полному торможению условных рефлексов. Видимо, двустороннее повреждение истощает симметричные компенсаторные механизмы.
При воздействиях на хвостатое ядро, помимо нарушений высшей нервной деятельности, отмечаются расстройства движения. Многие авторы отмечают, что у разных животных при двустороннем повреждении полосатого тела появляется безудержное стремление двигаться вперед, при одностороннем — возникают манежные движения.
Несмотря на большое функциональное сходство хвостатого ядра и скорлупы, имеется ряд функций, специфичных для последней. эволюционно скорлупа появляется раньше хвостатого ядра (ее зачатки есть уже у рыб).
Для скорлупы характерно участие в организации пищевого поведения: пищепоиска, пищенаправленности, пищезахвата и пищевладения; ряд трофических нарушений кожи, внутренних органов (например, гепатолентикулярная дегенерация) возникает при нарушениях функции скорлупы. Раздражения скорлупы приводят к изменениям дыхания, слюноотделения.
Как упоминалось ранее, раздражение хвостатого ядра тормозит условный рефлекс на всех этапах его реализации. В то же время раздражение хвостатого ядра препятствует угашению условного рефлекса, т. е. развитию торможения; животное перестает воспринимать новую обстановку. Учитывая, что стимуляция хвостатого ядра приводит к торможению условного рефлекса, следовало бы ожидать, что разрушение хвостатого ядра вызовет облегчение условнорефлекторной деятельности. Но оказалось, что разрушение хвостатого ядра также приводит к торможению условнорефлекторной деятельности. Видимо, функция хвостатого ядра не является просто тормозной, а заключается в корреляции и интеграции процессов оперативной памяти. Это подтверждается также тем, что на нейронах хвостатого ядра конвергирует информация различных сенсорных систем, так как большая часть этих нейронов полисенсорна.
№1.3. Функциональная организация нигро-стриопаллидарной системы
Стриопаллидарная система разделяется по функциональному значению и морфологическим особенностям на стриатум и паллидум (табл. 3).
Таблица 3. Функциональные и морфологические различия стриатума и паллидума
Никакая другая система не поддается в такой степени внешнему моделирующему влиянию гормонов и нейромедиаторов, как экстрапирамидная, так как функционально она входит в структуру лимбической системы. Именно поэтому энергетический настрой движений человека зависит от эмоций, и при поражении подкорковых ганглиев выявляются аффективные нарушения (насильственный смех и плач). Функции экстрапирамидной системы.
1. Регуляция мышечного тонуса в комплексе с другими структурами.
2. Регуляция темпа, ритма и пластики любого произвольного двигательного акта.
3. Обеспечение двигательного компонента в регуляции безусловных рефлексов (половой, оборонительный, старт-рефлекс и др.).
4. Обеспечение последовательности двигательного акта.
5. Обеспечение моторного компонента эмоциональной сферы.
6. Регуляция высокоспециализированных движений человека, которые достигли уровня автоматизмов.
№1.4. Симптомы поражения нигро-стриопаллидарной системы
Поражение паллидума: паллидарный синдром
Симптомокомплекс поражения бледного шара и черного вещества носит название паркинсонизма, акинетико-ригидного синдрома, амиостатического синдрома, гипертонически-гипокинетического синдрома. Он связан с функциональным дефицитом дофамина, с изменением влияния паллидонигральной системы на ретикулярную формацию и нарушением импульсации в корково-подкорково-стволовых нейронных кругах. Ретикулярная формация — стволовой «контролер-регулировщик» потока восходящих и нисходящих импульсов. При нарушении ее связей с черным веществом не препятствует прохождению к мышце избыточных тонических сигналов, вследствие чего развивается мышечная ригид- ность, поддерживаемая непрерывным потоком афферентных импульсов к стриопаллидарной системе. Возникает порочный круг: пораженная паллидарная система шлет бесконтрольные тонические сигналы, которые повышают мышечный тонус и усиливают поток импульсов обратной афферентации.
Основными симптомами поражения бледного шара являются:
• изменение мышечного тонуса по пластическому типу, феномен «зубчатого колеса» (сопротивление, испытываемое при исследовании тонуса, нарастает к концу движения), пластическая ригидность;
• поза восковой куклы, манекена — феномен каталепсии (при переходе из положения покоя в состояние движения больные часто застывают в неудобной позе);
• поза «просителя» — характерный внешний вид больных: туловище слегка согнуто, голова наклонена вперед (рис. 4.2), руки согнуты иприведены к туловищу, взгляд устремлен вперед, неподвижен;
• олигокинезия — бедность и маловыразительность движений; руки при ходьбе неподвижны, отсутствуют нормальные физиологические синкинезии;
• брадикинезия — замедленность движений; больные малоподвижны, инертны, скованы;
• паркинсоническое топтание на месте — затруднено начало двигательного акта, поэтому вначале каждого движения больной совершает несколько повторных движений, например, раскачивается или шагает на месте;
• иннерционое движение вперед — пропульсия, в сторону — латеропульсия, назад — ретропульсия; выведенный из состояния равновесия больной не может выровнять его автоматически;
• брадилалия — монотонная, замедленная, тихая речь;
• брадипсихия — замедление темпа мышления;
• микрография — почерк мелкий, нечеткий;
• акайрия — «вязкость» в общении, прилипчивость;
• «парадоксальные кинезии» — больные, целыми днями сидящие в кресле, в момент аффективных вспышек и эмоционального напряжения могут взбегать по лестнице, прыгать, танцевать;
• паркинсонический тремор покоя: чаще локализуется в пальцах кисти — феномен «катания пилюль», «счета монет», тремор головы — феномен «да-да, нет-нет, нет-да»; дрожание наблюдается в покое и уменьшается при произвольных движениях;
• положительные постуральные рефлексы: стопный феномен Вестфаля (при пассивном тыльном сгибании стопы возникает тоническое напряжение разгибателей стопы — она застывает в положении тыльного сгибания) и феномен голени (у больного, лежащего на животе с ногами, согнутыми в коленях под прямым углом, при дальнейшем пассивном сгибании голени она застывает в положении сгибания);
• нарушение ритма сон-бодрствование;
• вегетативные расстройства (сальное лицо, шелушение кожи, гиперсаливация).
Поражение полосатого тела: стриарный синдром
При поражении стриарной системы возникает дистонически- гиперкинетический синдром, обусловленный дефицитом тормозящего влияния стриатума на нижележащие двигательные центры, вследствие чего развиваются мышечная гипотония и избыточные непроизвольные движения (гиперкинезы).
Гиперкинезы — автоматические, чрезмерные движения, в которых участвуют отдельные части тела и конечности. Они возникают непроизвольно, исчезают во сне и усиливаются при произвольных движениях и волнении.
Атетоз — медленные, червеобразные, вычурные движения в дистальных отделах конечностей (в кистях и стопах) [рис. 4.3]. Атетоз в мышцах лица сопровождается выпячиванием губ, перекашиванием рта, гримасничаньем, прищелкиванием языком. Обычно атетоз связывают с поражением крупных клеток стриарной системы. Характерным его признаком является образование преходящих контрактур (spasmus mobilis), которые придают кисти и пальцам своеобразное положение.
Баллизм, гемибаллизм — крупные, размашистые, «бросковые» движения конечностей. Чаще всего баллизм затрагивает мышцы рук, вызывая движение в виде взмаха крыла птицы. Насильственные движения при гемибаллизме производятся с большой силой, их трудно прекратить. Возникновение гемибаллизма связывают с поражением льюисова тела, расположенного под зрительным бугром.
Хорея — быстрые сокращения различных групп мышц лица, туло- вища и конечностей. Гиперкинез неритмичен, некоординирован, распространяется на крупные мышцы дистальных и проксимальных отделов. Может напоминать произвольные движения, так как в процесс вовлекаются синергисты. Отмечаются нахмуривание бровей, лба, высовывание языка, порывистые, беспорядочные движения конечностей. Гиперкинез, охватывающий половину тела, называется гемихореей. Хорея возникает при поражении неостриатума и наблюдается при подкорковых дегенерациях, ревматическом поражении мозга, болезни Гентингтона.
В некоторых случаях хореические гиперкинезы сочетаются с атетозом (хореоатетоз). Хореоатетоз может наблюдаться у больных как постоянно, так и в виде приступов — пароксизмальный хореоатетоз. Описано несколько вариантов семейной формы пароксизмального хореоатетоза.
Дистония — насильственные сокращения мышц, приводящие к «выкручиванию», переразгибанию части тела. Выделяют спастическую кривошею (локальную дистонию мышц шеи), при которой голова повернута в сторону и наклонена к плечу (рис. 4.4). Возможны также непроизвольные наклоны головы вперед или назад. В начале заболевания напряжение мышц бывает преходящим, однако со временем оно становится постоянным, вследствие чего голова все время находится в неестественном положении. Спастическая кривошея нередко возникает в начальных стадиях торсионной дистонии в качестве локального ее проявления. Торсионная дистония является генерализованным вариантом гиперкинеза. Движения туловища носят вращательный, штопорообразный характер, сопровождаются гиперлордозом, сколиозом, вычурными позами. Торсионная дистония может прекращаться при различных компенсаторных приемах, например при обхвате руками шеи, усиленном повороте плеча и т.д. Выделяют также дистонию других групп мышц.
• Писчий спазм (графоспазм) — судорожное сокращение пальцев кисти, которое появляется во время письма.
• Профессиональные судороги — спазм мышц, участвующих в определенных профессиональных движениях. Наблюдаются у скрипачей, пианистов, гитаристов, машинисток и т.д.
• Лицевой гемиспазм — периодически повторяющиеся судороги мышц половины лица, иннервируемых лицевым нервом. Судороги сопровождаются появлением морщин на лбу, угол рта оттянут кнаружи и кверху, платизма напряжена.
• Лицевой параспазм Мейджа — периодически повторяющиеся симметричные судороги лицевых мышц. Параспазм часто возникает во время разговора, улыбки.
Блефароспазм — судорожные сокращения круговой мышцы глаза. Клинически блефароспазм проявляется частым миганием, возникает пароксизмально.
Икота — клонические судороги диафрагмы. Проявляется быстры- ми громкими вдыхательными движениями, обусловлена патологическим процессом в оболочках или в веществе мозга, интоксикацией.
Миоклонус — быстрый гиперкинез, который выглядит как вздра- гивание. Выделяют локальный миоклонус (например, конечности) и генерализованный. Следует также отличать неэпилептический миоклонус (гиперкинез) от эпилептического миоклонуса.
Тики — быстрые клонические подергивания ограниченной группы мышц, как правило, стереотипного характера, имитирующие произвольные движения. Чаще локализуются в мышцах лица и проявляются быстрым наморщиванием лба, поднятием бровей, миганием, высовыванием языка. Тик шейных мышц сопровождается поворотом головы в сторону, киванием. У детей тик нередко развивается как проявление невроза в результате образующегося патологического условного рефлекса, как подражание лицам, страдающим гиперкинезами. Тик лицевой мускулатуры может возникнуть при невралгии тройничного нерва. Тики могут вовлекать мышцы конечностей, туловища, диафрагмы. Особое место занимает генерализованный импульсивный тик — синдром де ля Туретта, при котором наблюдаются импульсивные подпрыгивания, приседания, гримасничанье, вокальные феномены в виде похрюкивания, вскриков, выкрикивания бранных слов (копролалия).
Дрожание (тремор) — стереотипный клонический ритмичный гиперкинез, преимущественно наблюдающийся в кистях рук, стопах; может также отмечаться дрожание туловища, головы. Дрожание — внешнее проявление нередко невидимого сокращения мышц в результате поражения сегментарных и надсегментарных двигательных структур, прежде всего стриопаллидарной системы и мозжечка. Амплитуда дрожания и его частота, длительность отдельных фаз могут быть различными в зависимости от механизма возникновения. Различают крупноразмашистый, «рубральный» тремор, возникающий при поражении красного ядра и захватывающий противоположную половину тела. Тремор может усиливаться или исчезать при движении. Интенционный тремор резко усиливается или возникает при выполнении целенаправленных движений, в частности при выполнении координаторных проб — пальце-носовой и пяточно-коленной. Статический тремор наблюдается в покое, при движениях не усиливается (напротив, может уменьшаться), характерен для паркинсонизма.
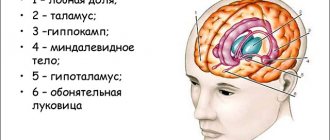
№2. Состав и функции лимбической системы
Лимбическая система представляет собой функциональное объединение структур мозга, участвующих в организации эмоционально-мотивационного поведения.
Что нужно знать о базальных ядрах
Большие полушария мозга человека снаружи представляют собой кору, образованную серым веществом, а внутри – подкорку из белого вещества. Базальные ядра (ганглии, узлы), которые также называют центральными, или подкорковыми – это сосредоточения серого вещества в белом веществе подкорки.
Базальные ганглии расположены в основании головного мозга, что и объясняет их название, снаружи от таламуса (зрительного бугра). Это парные образования, которые симметрично представлены в обоих полушариях мозга. При помощи нервных отростков они в двустороннем порядке взаимодействуют с различными областями центральной нервной системы.
Основная роль подкорковых узлов заключается в организации двигательной функции и различных аспектов высшей нервной деятельности. Патологии, которые возникают в их строении, влияют на работу других частей центральной нервной системы, вызывая проблемы с речью, координацией движений, памятью, рефлексами.
Особенности строения базальных узлов
Базальные ганглии находятся в лобных и частично височных долях конечного мозга. Это скопления тел нейронов, которые образуют группы серого вещества. Окружающее их белое вещество представлено отростками нервных клеток и формирует прослойки, разделяющие отдельные базальные ядра и прочие мозговые структурно-функциональные элементы.
К базальным узлам причисляются:
- полосатое тело;
- ограда;
- миндалевидное тело.
На анатомических срезах полосатое тело выглядит как перемежающиеся слои серого и белого вещества. В его составе выделяют хвостатое и чечевицеобразное ядра. Первое расположено кпереди от зрительного бугра. Истончаясь, хвостатое ядро переходит в миндалевидное тело. Чечевицеобразное ядро находится латеральнее зрительного бугра и хвостатого ядра. Оно соединяется с ними тонкими перемычками нейронов.
Ограда – это узкая полоска нейронов. Она расположена между чечевицеобразным ядром и островковой корой мозга. От этих структур ее отделяют тонкие слои белого вещества. Миндалевидное тело по форме напоминает миндалину и находится в височных долях конечного мозга. В его составе различают несколько самостоятельных элементов.
Данная классификация построена на особенностях строения и расположения ганглиев на анатомическом срезе мозга. Существует также функциональная классификация, согласно которой ученые причисляют к базальным узлам только полосатое тело и некоторые ганглии промежуточного и среднего мозга. Эти структуры в комплексе обеспечивают двигательные функции человека и отдельные аспекты поведения, отвечающие за мотивацию.