У полушарий мозга
Кора – это структура мозга и слой серого вещества, имеющий толщину от 1,3 до 4,5 мм. Он располагается по периферии каждого из полушарий, покрывая их. Наибольшая его толщина находится в верхних частях постцентральной и предцентральной извилин, а также предцентральной доли (дольки).
Кора полушарий мозга играет важнейшую роль при осуществлении высшей психической (нервной) деятельности всего организма. Она составляет более 80 % от всей его массы. У человека в среднем она равняется примерно 44 % от общего объема каждого из полушарий.
- Отделы головного мозга – их строение и функции
- Клетки головного мозга
- Особенности строения
- Патологии и признаки поражения отделов
- Клещевой энцефалит
- Строение коры головного мозга
- Строение
- Внутри черепа есть мост
- Белое вещество
- Изучение развития головного мозга
Кора полушарий разделяется на четыре типа:
- Палеокортекс (древняя).
- Архикортекс (старая).
- Неокортекс (новая).
- Промежуточная (состоит из промежуточных частей палеокортекса и архикортекса).
Эта оболочка покрывает большой головной мозг полностью и образует довольно большое количество борозд, которые отличаются глубиной и протяженностью. Между ними располагаются различные по величине извилины.
На сегодняшний день кора головного мозга и все остальные его части изучены довольно глубоко. Известно, какие области мозга отвечают за те или иные процессы в организме. Однако все же еще существует вопросы, не имеющие ответов.
Как видно из сказанного, кора – это термин, имеющий большое количество значений, которые затрагивают различные сферы жизни человека.
Лечение повреждения лобной доли
Лечение повреждения лобной доли направлено на устранение причины травмы. Врач может прописать препараты при инфекции, провести операцию или назначить средства, чтобы уменьшить риск развития инсульта.
В зависимости от причины травмы назначают лечение, которое может помочь. Например, при фронтальном повреждении после инсульта необходимо перейти к здоровой диете и физической активности, чтобы уменьшить риск инсульта в будущем.
Препараты могут быть полезны для людей, у которых нарушены внимание и мотивация.
Лечение повреждений лобной доли требует постоянного ухода. Восстановление после травмы часто является длительным процессом. Прогресс может прийти внезапно, и его невозможно полностью предсказать. Восстановление тесно связано с поддерживающей терапией и здоровым образом жизни.
Отделы головного мозга – их строение и функции
Центр нервной системы представляет собой и единый «управляющий блок», и группу слаженно работающих отдельных структур. Основные отделы головного мозга, задний, средний и передний, выполняют определенные функции, поэтому отличаются по строению и энергозатратности. Даже клетки, из которых состоит каждая область органа, имеют свои особенности строения.
Задний отдел головного мозга
Указанная область включает две структурные единицы:
- Варолиев мост.
Продолжение продолговатого мозга, с другой стороны граничит со средним отделом. Задачей моста является передача нервных сигналов. Через него проходят все пути (восходящие и нисходящие), которые связывают спинной мозг с передним, мозжечком и другими частями органа. Варолиев мост содержит ядра лицевого, тройничного и отводящего и нервов. Он отвечает за множество рефлексов – глазные, моргание, мочеиспускание, моторику кишечника и другие.
Мозжечок
. Располагается под затылочными долями больших полушарий. Строение головного мозга позволяет мозжечку сообщаться с соседними отделами посредством трех пар «ножек». Данная структура занимает всего 10% площади, но содержит более половины всех имеющихся нейронов. Главные функции мозжечка – вегетативная регуляция, координация движений, сохранение равновесия и мышечного тонуса. Дополнительно он поддерживает работу желудочно-кишечного тракта, сердечно-сосудистой системы. В последние 10 лет исследования показали, что мозжечок участвует и в высшей нервной деятельности – памяти, мышлении, накоплении опыта.
Продолговатый отдел головного мозга
Альтернативные названия – миелэнцефалон и луковица. Отдел представляет собой прямое продолжение спинного мозга. Он располагается у основания черепа в его задней части. Задачи луковицы – контроль дыхания и процессов кровообращения. Строение головного мозга и функции в данном случае находятся в тесной зависимости, поэтому при повреждении миелэнцефалона его работа не компенсируется другими отделами, и мгновенно наступает смерть. Дополнительно луковица помогает поддерживать равновесие, обмен веществ, координацию и мышечную деятельность, связь с органами чувств.
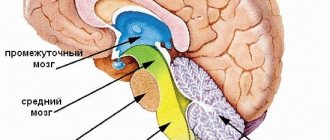
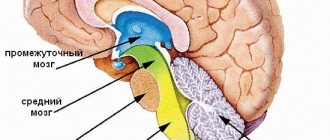
Средний отдел головного мозга
Одна из самых маленьких структур, около 2-х см в диаметре. Располагается между задним и промежуточным мозгом. Строение включает следующие части:
- крыша (зеленый);
- среднемозговая область покрышки (красный);
- Сильвиев водопровод (черный);
- ножки (голубой).
Небольшой размер не мешает этому отделу выполнять важнейшие функции головного мозга, которые часто ошибочно приписывают полушариям:
- общий уровень возбуждения центральной нервной системы;
- слух;
- зрение;
- концентрация внимания;
- оборонительные, ориентировочные и защитные рефлексы;
- контроль движений;
- поддержание нормальной температуры тела.
В описываемом отделе происходит выработка дофамина, поэтому средний мозг регулирует:
- эмоциональную активность;
- репродуктивное поведение и возбудимость;
- уровень мотивации;
- развитие привычек и пристрастий, включая зависимости;
- болевую чувствительность;
- циклы сна и бодрствования.
Передний мозг
Строение этой части центральной нервной системы включает диэнцефалон и большие полушария. Другие названия – промежуточный и конечный мозг. Диэнцефалон представляет собой «подложку» полушарий, но имеет сложное строение:
- Таламический мозг.
Включает таламус, эпиталамус и субталамус. - Гипоталамический отдел (гипоталамо-гипофизарная система).
К нему относится задняя доля гипофиза и гипоталамус.
Промежуточный мозг выполняет массу важных функций в организме. Он регулирует:
- температуру тела;
- режимы сна и бодрствования;
- жажду и голод;
- ощущение насыщения;
- нейрогуморальные процессы;
- работу желез внутренней секреции;
- выработку гормонов.
Конечный мозг состоит из правого и левого полушарий. Они соединяются друг с другом поперечными нервными пучками (комиссурами). Главным «мостом» между полушариями считается пластина мозолистого тела. В популярной психологии пропагандируется утверждение, что строение конечного головного мозга предполагает разрозненную работу его половинок. Одна отвечает за логику и рациональность, другая – за творчество и фантазию.
Современные научные исследования опровергают теорию асимметрии. Оба полушария головного мозга параллельно принимают участие в высшей нервной деятельности. Распределение задач неравномерное, но отличия несущественные. Функции конечного мозга:
регуляция движений; зрение; речь; кожно-мышечная чувствительность; обоняние; слух; вкусовые ощущения; мышление; логика; внимание; память; обучение; формирование эмоций и другие.
Общие сведения
Образован из переднего конца первичной нервной трубки. В эмбриогенезе делится на 2 части, одна из которых порождает конечный мозг, вторая – промежуточный.
Согласно модели Александра Лурии, состоит из 3-х блоков:
- Блок регуляции уровни активности мозга. Обеспечивает осуществление определённых видов деятельности. Отвечает за эмоциональное подкрепление активности на основе прогнозирования ее результатов (успех – неудача).
- Блок приёма, переработки и хранения входящей информации. Участвует в формировании представления о способах реализации деятельности.
- Блок программирования, регуляции и контроля за организацией психической деятельности. Сравнивает полученный результат с исходным намерением.
Передний мозг принимает участие в работе всех блоков. На основе обработки информации он управляет поведением. Администратор высших психологических функций: восприятия, памяти, воображения, мышления, речи.
Клетки головного мозга
Мозг состоит из двух типов клеток: нервных клеток (нейронов) и глиальных клеток.
Нервная клетка
Существует множество размеров и форм нейронов, но все они состоят из тела клетки, дендритов и аксона. Нейрон передает информацию посредством электрических и химических сигналов. Попробуйте представить себе электрическую проводку в вашем доме. Электрическая цепь состоит из многочисленных проводов, соединенных таким образом, что при включении выключателя света будет светиться лампочка. Возбужденный нейрон будет передавать свою энергию нейронам, находящимся в непосредственной близости от него.
Важно Причины, симптоматика и разновидности тахилалии
Нейроны передают свою энергию, или “говорят”, друг другу через крошечный промежуток, называемый синапсом (рис. 12). У нейрона есть много рук, называемых дендритами, которые действуют как антенны, собирающие сообщения от других нервных клеток. Эти сообщения передаются в тело ячейки, которое определяет, следует ли передавать сообщение вместе.
Важные сообщения передаются в конец аксона, где мешочки, содержащие нейротрансмиттеры, открываются в синапс. Молекулы нейротрансмиттера пересекают синапс и помещаются в специальные рецепторы на принимающей нервной клетке. Это стимулирует клетку передавать сообщение.
Рисунок 12. Нервные клетки состоят из тела клетки, дендритов и аксона. Нейроны взаимодействуют друг с другом, обмениваясь нейротрансмиттерами через крошечный промежуток, называемый синапсом.
Глиальные клетки
Glia (греческое слово, означающее клей) — это клетки головного мозга, которые обеспечивают нейроны питанием, защитой и структурной поддержкой. Существует примерно в 10-50 раз больше glia, чем нервные клетки и являются наиболее распространенным типом клеток, участвующих в опухолях головного мозга.
Особенности строения
Лобные доли – отдел головного мозга, который несет ответственность за коммуникативные навыки (умение разговаривать, мыслить, запоминать). Анатомическое строение лобной доли предполагает четкое ограничение со стороны височных областей боковыми бороздами. С фронтальной стороны лобная часть отделяется от теменного участка центральной бороздой. Нижний край отдела ограничивает сильвиевая борозда.
Основные извилины, находящиеся в лобной доле, включают вертикальную прецентральную и верхнюю, среднюю, нижнюю, пролегающие горизонтально. Извилины разделены бороздами. В период бодрствования нейроны и нейромедиаторы в этой области более активны, чем во время сна.
Участок префронтальной коры функционально связан и активно взаимодействует с отделами лимбической нервной системы – мозговыми структурами, находящимися по обеим сторонам таламуса. Лимбическая система задействована в регуляции обоняния, работы внутренних органов, проявления эмоций, режима сна и бодрствования, запоминания.
Гипоталамус, входящий в лимбическую систему, управляет при помощи гормонов вегетативной нервной системой, благодаря чему человек чувствует жажду и голод, просыпается и засыпает в соответствии с заложенными природой биологическими часами, испытывает половое влечение к представителям противоположного пола. Гиппокамп участвует в процессе формирования базы долговременной памяти. Эта структура ответственна за восприятие, анализ и хранение информации пространственного плана.
Лобная доля считается корковым участком лимбической системы. В лобной части мозга находятся ассоциативные зоны, где происходит обработка и сравнение поступающей извне информации с данными, которые хранятся в памяти. Анатомия этой мозговой структуры предполагает деление на отделы:
- Моторный.
- Премоторный.
- Дорсолатеральный префронтальный.
- Медиальный префронтальный.
- Орбитофронтальный. Является пересечением нейронных путей, по которым передается информация в ассоциативные зоны коры – место тесного взаимодействия префронтальной и лимбической системы.
Префронтальная область регулирует сложные формы и модели поведения, обеспечивая взаимосвязь мотивационных, эмоциональных и мыслительных процессов. Отдел участвует в оценке окружающей обстановки, с учетом обстоятельств формирует порядок планируемых действий, анализирует последствия намеченных поступков, вырабатывает типичные для конкретных ситуаций модели поведения.
Функции коры
Кора головного мозга человека отвечает за контроль над множеством функций, которые используются в разных аспектах жизнедеятельности организма человека. Ее толщина составляет около 3-4 мм, а объем довольно внушительный за счет наличия связующих с центральной нервной системой каналов. Как по электросети происходит восприятие, обработка информации, прием решений с помощью нервных клеток с отростками.
Внутри коры головного мозга вырабатываются различные электросигналы (тип которых зависит от текущего состояния человека). Активность этих электрических сигналов зависит от самочувствия человека. Технически электросигналы такого типа описываются с помощью показателей частоты и амплитуды. Большее количество связей и нейронов локализовано в местах, которые несут ответственность за обеспечение наиболее сложных процессов. При этом кора головного мозга продолжает активно развиваться в течение всей жизни человека (по крайней мере, до того момента, пока развивается его интеллект).
В процессе обработки информации, поступающей в мозг, в коре формируются реакции (психические, поведенческие, физиологические и т.д.).
Наиболее важными функциями коры мозга являются:
- Взаимодействие внутренних органов и систем с окружающей средой, а также друг с другом, правильное течение обменных процессов внутри организма.
- Качественный прием и обработка получаемой информации извне, осознание полученной информации за счет протекания процессов мышления. Высокая чувствительность к любой получаемой информации достигается за счет большого количества нервных клеток с отростками.
- Поддержка беспрерывной взаимосвязи между различными органами, тканями, структурами и системами организма.
- Формирование и правильная работа сознания человека, течение творческого и интеллектуального мышления.
- Осуществление контроля над активностью речевого центра и процессами, связанными с разными психическими и эмоциональными ситуациями.
- Взаимодействие со спинным мозгом и другими системами и органами человеческого организма.
Кора головного мозга в своей структуре имеет передние (лобные) отделы гемисфер, которые на данный момент современной наукой изучены в наименьшей степени. Об этих участках известно, что они практически невосприимчивы к внешнему воздействию. Например, если на эти отделы воздействовать с помощью внешних электрических импульсов, они не будут давать никакой реакции.
Некоторые ученые уверены, что передние отделы больших полушарий отвечают за самосознание человека, за его специфичные особенности характера. Известен тот факт, что люди, у которых передние отделы поражены в той или иной степени, испытывают определенные сложности с социализацией, они практически не уделяют внимания своему внешнему виду, им не интересна трудовая деятельность, не интересует мнение окружающих.
С точки зрения физиологии, значение каждого отдела больших полушарий сложно переоценить. Даже тех, которые на данный момент до конца не изучены.
Патологии и признаки поражения отделов
Обширное поражение медиальных участков лобной доли провоцирует развитие абулии, которая проявляется замедленными реакциями, равнодушием, безучастностью к происходящему. При повреждении участка префронтальной орбитальной коры у пациента наблюдается отсутствие критической оценки собственного поведения, эмоциональная лабильность.
Двусторонняя травма в лобной области сопровождается признаками: ажитация, беспокойное поведение, навязчивость, многословие. Аномальное поведение является признаком деменции, которая развивается на фоне дегенеративных процессов, затрагивающих лобные доли. Повреждение мозгового вещества двигательной коры вызывает гемипарез или мышечную слабость.
Нарушения развиваются на стороне, противоположной местоположению патологического очага в мозге. Повреждение зрительного участка в одном полушарии приводит к развитию двухсторонней слепоты в половине поля обзора. Поражение поля 19 ассоциируется со зрительной агнозией – нарушением зрительного восприятия. Пациент видит предмет, но не может его распознать.
Информация, которая поступает через зрительный анализатор, не обрабатывается или обрабатывается неправильно, что приводит к невозможности различать знакомые предметы, лица людей. У таких больных нарушается цветовое восприятие – они не различают оттенки.
Повреждение поля 22 приводит к развитию музыкальной глухоты (нарушение восприятия музыкальных произведений), появлению слуховых галлюцинаций, нарушению реакций, ориентированных на слуховые раздражители. Поражение поля 41 сопровождается развитием корковой глухоты (невозможность восприятия звуковых сигналов).
Поражение поля 34 сопровождается нарушением восприятия запахов, в том числе обонятельными галлюцинациями. Патологические структурные изменения в нервной ткани поля 39 приводят к неспособности читать и писать. При повреждении ткани поля 37 человек не помнит названия предметов.
Зоны мозга разделяются на сенсорные и двигательные, а также ассоциативные – причем все участки взаимодействуют между собой. Каждый отдел наделен определенными функциями, которые в совокупности обуславливают высшую психическую и сложную двигательную деятельность.
PsyAndNeuro.ru
История изучения психомоторных нарушений с момента своего начала в первых годах XIX-го века представляет собой сложную и запутанную последовательность сменявших друг друга парадигм и концепций. Все началось с Wilhelm Griesinger, впервые представившего концепцию «психомоторных нарушений». Однако его последователи объединяли под этим термином немного разное: Emil Kraepelin – сенсомоторные нарушения, Karl Kahlbaum и August Hoch – психомоторную концепцию кататонии, Gilles de la Tourette – непроизвольные движения, Jean-Martine Charcot и Pierre Marie – двигательные симптомы истерического паралича. Затем Carl Wernicke и Karl Kleist описали так называемые «двигательные» и «циклоидные» психозы как два обособленных эпизодических состояния со своими психомоторными проявлениями. Однако за удивительно малое время – с появлением первых антипсихотиков в 1950-х – концепция подлинно психомоторных нарушений была практически забыта, и «все дороги стали вести» к исключительно моторным нарушениям. Только в 1960-х Karl Leonhard (ученик Karl Kleist) начал возвращать внимание к собственно психомоторным нарушениям.
Несмотря на то, что психомоторные нарушения являются одними из ключевых особенностей таких состояний, как большое депрессивное расстройство (БДР), биполярное аффективное расстройство (БАР) и шизофрения, в общепринятых современных диагностических классификациях (МКБ-10 и DSM-5) делается упор на когнитивные, аффективные и социальные симптомы, но не на психомоторные. В дименсиональной классификации RDoC двигательные симптомы психических заболеваний все же рассматриваются, однако в ней все же делается упор на связанные исключительно с двигательной функцией изменения дофаминергических кортико-стриато-паллидо-таламо-кортикальных моторных путей. В отличие от данных подходов, в нашей работе мы поставили цель возродить концепцию по-настоящему «психо-моторного» в буквальном смысле этого слова, показав специфические психомоторные (а не просто моторные) механизмы.
Что конкретно мы понимаем под термином «психомоторный»? Очевидно, что данное слово состоит из двух частей: «психо» и «моторный». В отличии от редуцирования первого компонента, т.е. «психики», в угоду второму, т.е. «моторному», как это часто происходит, мы постарались непосредственно оценить прямую связь между психическими и двигательными функциями и рассмотреть лежащие в основе данной связи нейробиологические механизмы. На нейрональном уровне психомоторные механизмы обуславливают то, как первичные моторные функции (т.е. дофаминергические субкортикально-кортикальные пути) модулируются немоторными функциями, то есть когнитивными и эмоциональными системами. С психологической точки зрения психомоторные механизмы обуславливают движения тела как результат некой психической активности, что необходимо для взаимодействия с окружением и объектами. Этот взгляд соотносится с первоначальным пониманием «психомоторного», подчеркивающим само взаимодействие аффективных, когнитивных и моторных функций. Как указано в названии нашей работы, мы покажем патофизиологические механизмы, посредством которых аффективные и когнитивные изменения, не связанные с двигательными зонами тем или иным образом (т.е., разными дорогами) приводят к дисфункции моторной коры и, соответственно, к появлению психомоторной симптоматики.
В нашей работе мы стремились возродить традиционное понимание психомоторных феноменов как чего-то большего, чем просто двигательные нарушения. Ведь согласно данным последних исследований, нейрональные и биохимические механизмы, лежащие в основе психомторных нарушений, затрагивают гораздо большее число систем головного мозга, чем одна лишь дофаминергическая базально-таламо-кортикальная моторная петля. Мы построили свой обзор не по традиционной схеме, отталкиваясь от нозологий, а, скорее следуя концепции RDoC, сгруппировали информацию по различным психомоторным синдромам.
В данной работе мы приводим результаты МРТ-исследований БДР, БАР и шизофрении, посвященных поиску нейробиологических коррелятов психомоторных феноменов. В начале мы фокусируемся на том, как именно дофаминергическая субкортикально-кортикальная моторная система модулируется другими нейромедиаторными системами, в частности, серотониновыми путями (субкортикально-кортикальная модуляция). Затем мы описываем, как сенсомоторная сеть (sensorimotor network, SMN) и соответствующие двигательные функции модулируются другими, немоторными, сетями, в частности, сетью пассивного режима работы мозга (default mode network, DMN). Также мы описываем, как общая активность коры влияет на локальную активность моторных отделов коры (т.е. кортико-кортикальную модуляцию).
Изменения в ядре шва модулируют активность субкортикально-кортикальных моторных путей
Одним из важных отделов ствола мозга, влияющих на дофаминергическую моторную сеть, является серотонинергическое ядро шва (raphe nucleus, RN). Так как в настоящее время опубликовано достаточно небольшое число работ, посвященных прямому влиянию ядра шва на моторную кору, в данном разделе мы также приводим данные фМРТ исследований самого ядра шва.
Han et al. в своем исследовании изучали функциональную связность в состоянии покоя (resting-state functional connectivity, rsFC (под этим термином понимают временную корреляцию изменения активности различных зон головного мозга; то есть, две зоны считаются более функционально связанными, если паттерны их возбуждения более синхронизированы, при условии, что обе эти зоны являются частями одной функциональной сети – прим. пер.)) ядра шва с остальными зонами мозга у пациентов с БАР и БДР. Оказалось, что при БАР и при БДР наблюдаются противоположные паттерны rsFC ядра шва с остальными субкортикальными регионами, в частности, с таламусом и скорлупой. Кроме того, при биполярном расстройстве rsFC гипоталамуса была повышена, а при депрессии, наоборот, снижена. Эти находки были подкреплены результатами исследования Anand et al., в котором было обнаружено значимое снижение rsFC ядра шва с префронтальной корой и средней поясной извилиной при БДР. Кроме того, показатель rsFC ядра шва с гиппокампом и миндалиной коррелировал с выраженностью депрессивных симптомов. Наконец, Wohlschlager et al. пошли дальше исследования rsFC и изучили спектральные характеристики инфрамедленных колебаний (0,01-0,1 Гц) активности ядра шва и вентральной зоны покрышки (ventral tegmental area, VTA) у пациентов с БДР, не получавших терапию. Было обнаружено, что в обеих зонах происходит существенное усиление более низкочастотной активности, что также прямо коррелировало с выраженностью депрессивной симптоматики.
Все эти данные свидетельствуют, что при депрессии имеют место определенные изменения в самом ядре шва, а также в его связях с субкортикальными моторными зонами (таламусом и базальными ганглиями) и различными немоторными зонами коры. Однако в настоящее время крайне мало исследований, посвященных возможным функциональным изменениям ядра шва и его rsFC, а также серотонинергической дисфункции, при шизофрении. Тем не менее, в указанных выше исследованиях предполагалось, что аномалии функциональных связей ядра шва (как центральной структуры серотонинергической системы) с другими субкортикальными и кортикальными регионами могут являться трансдиагностическими феноменами. Все это оставляет открытым вопрос о связи этих аномалий с моторной функцией и особенно с психомоторными симптомами.
Модуляция дофаминовых субкортикально-кортикальных моторных путей ядром шва и серотонином в здоровом мозге
Влияет ли серотониновое ядро шва на дофаминергические субкортикально-кортикальные пути и моторные функции? Conio et al. на основе данных фармакологических, структурных и функциональных МРТ-исследований продемонстрировали наличие сложных взаимодействий между серотонином и дофамином, а также влияние этих взаимодействий на активность головного мозга. В частности, было показано, что дофаминергическая черная субстанция соединена, в основном, с сенсомоторной сетью (а VTA – с сетью салиентности (salience network, SAN). Однако серотонинергическое ядро шва имеет связи как с сенсомоторной сетью, так и с сетью пассивного режима работы мозга. Помимо различных связей дофаминовых и серотониновых ядер ствола мозга, активность дофаминовых путей приводит к усилению активности SMN, а активность серотонинергических путей, наоборот, к уменьшению активности SMN и усилению активности DMN. Более того, rsFC черной субстанции прямо коррелировали с активностью SMN, а rsFC ядра шва коррелировали с активностью SMN обратным образом. Martino et al. в своем исследовании показали, что у здоровых испытуемых rsFC между таламусом и SMN модулируются функциональными связями как от ядра шва, так и от черной субстанции, но противоположным образом. Иными словами, связи от черной субстанции способствуют большей синхронизации активности между таламусом и SMN, а связи от ядра шва, наоборот, уменьшают эту синхронизацию. Таким образом, на основании этих данных можно утверждать, что серотонин ядра шва и дофамин черной субстанции диаметрально противоположным образом влияют на активность моторных зон коры, в частности, на SMN. Дофаминовые сигналы усиливают активность SMN и ее синхронизацию с активностью таламуса, а серотониновые сигналы, наоборот, приводят к «антикорреляции» SMN и таламуса и снижению активности SMN.
Таким образом, отвечая на вопрос, поставленный в начале раздела, можно утверждать, что ядро шва и серотонин действительно модулируют дофаминовые субкортикально-кортикальные пути. Соответственно, дофаминовые пути от черной субстанции и серотониновые пути от ядра шва можно расценивать не как первично моторные пути сами по себе, а скорее как модуляторы психомоторных функций.
Модуляция дофаминовых субкортикально-кортикальных моторных путей ядром шва и серотонином при депрессии, БАР и шизофрении
Основываясь на данных, полученных при исследовании здоровых испытуемых, можно предположить, что психомоторное торможение, в основе которого лежит сниженная активность SMN, должно быть обусловлено уменьшением rsFC черной субстанции и/или увеличением rsFC ядра шва. Подобные феномены действительно наблюдаются у пациентов с психомоторным торможением в депрессивной фазе БАР. Для таких пациентов характерно уменьшение rsFC таламуса и SMN и черной субстанции с таламусом/базальными ганглиями, а также конкурентное уменьшение rsFC ядра шва с базальными ганглиями/таламусом. Вместе эти паттерны способствуют «отсоединению», то есть, рассинхронизации активности таламуса и SMN, с последующим торможением активности SMN и возникновением психомоторного торможения. Эти находки согласуются с результатами исследования Yin et al., в котором было продемонстрировано снижение кровотока в моторной коре при психомоторном торможении у пациентов с БДР.
Для пациентов с манией, наоборот, характерны практически диаметрально противоположные изменения. У данных пациентов наблюдалось уменьшение rsFC ядра шва с таламусом/базальными ганглиями, а также, в отличие от пациентов с депрессией, rsFC черной субстанции была сохранна. Все вместе это приводит к увеличению rsFC таламуса с SMN, усилению активности SMN и последующей психомоторной ажитации.
Таким образом, данные этих исследований свидетельствуют о наличии реципрокного баланса между дофаминергическими связями базальных ганглиев/таламуса с SMN с одной стороны и серотонинегической модуляцией этих связей ядром шва с другой стороны. Уменьшение активности серотонинергических путей от ядра шва до базальных ганглиев приводит к большей «связанности» талаумса c SMN, что приводит к психомоторному возбуждению. Уменьшение же активности дофаминергических путей от черной субстанции до базальных ганглиев, напротив, уменьшает «связанность» таламуса с SMN и приводит к возникновению психомоторного торможения. Если эти два плеча модуляции связей таламус-SMN, дофаминергическое и серотонинергическое, уравновешены, то психомоторная функция «нормальная», то есть, не ускорена и не замедлена. Если же этот баланс нарушается, то таламус и SMN становятся либо чрезмерно, либо недостаточно функционально связанны, что приводит либо к психомоторному возбуждению (например, при мании), либо к психомоторному торможению (при депрессии). Подробнее см. Рис. 1А.
Рис. 1. Биохимическая и нейрональная модуляция субкортикально-кортикальных и кортико-кортикальных механизмов психомоторных феноменов. А. Модуляция дофаминергической субкортикально-кортикальной моторной цепи ядром шва и серотонином. B. Модуляция сенсомоторной сети (SMN) сетью пассивного режима работы мозга (DMN). С. Модуляция отображения общей активности в SMN. SN – черная субстанция, RN – ядро шва, Thal – таламус.
Модуляция дофаминовой субкортикально-кортикальной моторной петли ядром шва/серотонином и другими нейромедиаторными системами – дименсиональный и транснозологический подход
Следующий вопрос, на который необходимо ответить, звучит следующим образом: можно ли соотнести серотониновую модуляцию моторной петли с различными диагностическими категориями, или, формулируя иначе, можно ли ее соотнести с различными степенями психомоторных нарушений? Своеобразной «лакмусовой бумажкой», способной помочь ответить на данные вопросы, является феномен психомоторного возбуждения при депрессии. Martino et al. отдельно исследовали пациентов в депрессивной фазе БАР с психомоторным возбуждением и сравнивали их с соответствующими пациентами с психомоторным торможением. Оказалось, что у депрессивных пациентов с психомоторным возбуждением наблюдается паттерн, схожий с тем, что имеет место при мании: при обоих состояниях наблюдается усиление rsFC таламуса с SMN, что, соответственно, приводит к усилению активности SMN и психомоторному возбуждению.
Еще одно доказательство дименсиональной и транснозологической природы двусторонней «от ядра шва и от черной субстанции) модуляции активности субкортикально-кортикальной моторной петли следует из результатов недавнего исследования шизофрении. Авторы изучали две группы пациентов с шизофренией – с психомоторным возбуждением и с психомоторным торможением – и сравнивали их соответственно с группами пациентов с БАР в фазе мании (и с психомоторным возбуждением) и в фазе депрессии (с сопутствующим психомоторным торможением). Оказалось, что для пациентов, страдающих шизофренией с психомоторным торможением, характерен почти тот же паттерн изменений таламо-сенсомоторной rsFC, что и для пациентов, страдающих депрессией с психомоторным торможением, то есть, снижение rsFC таламуса с SMN и сопутствующее уменьшение rsFC ядра шва и черной субстанции с базальными ганглиями и таламусом. Аналогичное совпадение наблюдалось и у пациентов, страдающих шизофренией с психомоторным возбуждением. Наблюдаемые у таких пациентов усиление rsFC таламуса c SMN и уменьшение rsFC только ядра шва с базальными ганглиями и таламусом аналогичны соответствующим изменениям у пациентов в маниакальной фазе БАР с психомоторным возбуждением.
Хрестомайтиным примером психомоторного синдрома является кататония, являющаяся по-настоящему трансдиагностическим феноменом. Кататония может встречаться при шизофрении, БАР, БДР и при других заболеваниях. Развитие кататонии является результатом дисфункции сразу нескольких нейромедиаторных систем. Действительно, кататония, в первую очередь, проявляется двигательными нарушениями, связанными с изменениями активности сенсомоторных субкортикально-кортикальных зон. Однако кататония также характеризуется аффективными нарушениями, связанными с изменениями в немоторных зонах коры (например, с нарушениями функционирования лобно-теменных связей), что подчеркивает истинно психомоторную природу этого феномена. Соответственно, наряду с гипоактивацией дофаминергических рецепторов, при кататонии наблюдаются нарушения в других нейромедиаторных системах: гипоактивация серотониновых (5-HT2A) рецепторов, дисбаланс между активностью ГАМКА (гипоактивация) и ГАМКВ (гиперактивация) рецепторов и, вероятно, также гиперактивация глутаматных NMDA-рецепторов. С одной стороны, лоразепам и золпидем (аллостерические модуляторы ГАМКА-рецепторов) усиливают возбудимость ГАМК-ергических ингибиторных сетей в моторной коре и, таким образом, способствуют облегчению симптомов кататонии. С другой стороны, баклофен и вальпроевая кислота могут усиливать активность ГАМКв- и NMDA-рецепторов и, наоборот, усилить выраженность кататонии. Тем не менее, все же существуют некоторые данные о положительном влиянии вальпроевой кислоты, топирамата и карбамазепина на аффективные кататонические симптомы (предположительно, усиливая восприимчивость ГАМК и NMDA). Наконец, клозапин (антагонист 5-HT2A-рецепторов и агонист ГАМКB-рецепторов) компенсирует серотонинергическую гипоактивацию дисбаланс активности ГАМКA-B и, таким образом, согласно некоторым исследованиям, может уменьшать выраженность кататонии.
Другим примером транснозологического психомоторного синдрома служит паркинсонизм (сочетание тремора, ригидности и брадикинезии), являющийся, главным образом, следствием болезни Паркинсона, первично нейродегенеративной по своей природе. Однако явления паркинсонизма могут быть ассоциированы и с другими заболеваниями (например, паркинсонизм может наблюдаться у больных шизофренией). Болезнь Паркинсона, в первую очередь, характеризуется дегенерацией дофаминергических клеток черной субстанции и стриатума, однако было обнаружено, что при данном заболевании также нарушается активность серотонинергической системы. Так, при болезни Паркинсона имеет место снижение количества серотониновых транспортеров в ядре шва, что прямым образом ассоциировано с тяжестью тремора. Кроме того, при болезни Паркинсона затронуты также другие нейромедиаторные системы. Известно, что модуляторы ГАМКергической системы (золпидем и клоназепам), амантадин, антихолинергические препараты и клозапин могут улучшать двигательную симптоматику болезни Паркинсона. У пациентов с шизофренией, страдающих паркинсонизмом, наблюдаются характерные изменения объема серого вещества и активности фронто-таламических/мозжечковых и кортикальных соматосенсорных сетей, не свойственные пациентам без паркинсонизма. Эти факты дают возможность предположить существование восходящей модуляции моторных отделов коры как центрального нейронального механизма возникновения паркинсонизма при шизофрении.
Наконец, психомоторные нарушения могут наблюдаться и при других заболеваниях, в частности, при аутизме. Стоит отметить, что имеются данные об ассоциированных с данным расстройством изменениях активности дофаминергической, серотонинергической (повышенный синтез серотонина), ГАМКергической и глутаматергической систем в моторной и соматосенсорной коре, а также в стриатуме. Соответственно, блокаторы дофаминовых рецепторов, ингибиторы обратного захвата серотонина, мемантин, вальпроевая кислота, арбаклофен и акампросат в некоторых исследованиях демонстрируют эффективность в уменьшении выраженности таких психомоторных симптомов аутизма, как стереотипии, импульсивность и раздражительность.
Все эти результаты ясно демонстрируют, что модуляция ядром шва и серотонином субкортикально-кортикальной моторной петли представляют собой явление, простирающееся за рамки каких-то конкретных нозологических категорий. Она соответствует скорее направленности и выраженности психомоторных нарушений, а не какому-то конкретному диагнозу, являясь, таким образом, по-настоящему дименсиональным и транснозологическим феноменом.
Модуляция моторной петли другими корковыми сетями
Кора головного мозга включает в себя различные нейронные сети: сеть пассивного режима работы мозга (DMN), сенсорные сети (зрительная, слуховая, соматосенсорная), сенсомоторная сеть (SMN), фронтопариетальная сеть, сеть салиентности, сети внимания и многие другие. Недавние исследования показали, что все эти сети функционально взаимосвязанны. К примеру, DMN и фронтопариетальная сеть противоположности по своей активности: функциональное усиление активности одной прямо приводит к угнетению активности другой и наоборот. Что особенно интересно, SMN также, по-видимому, находится в реципрокных отношениях с другими сетями, в частности, с DMN и сенсорными сетями. В исследовании Martino et al. было продемонстрировано, что увеличение нейрональной вариабельности (вариабельность амплитуды сигнала от определенной популяции нейронов; считается, что чем выше нейрональная вариабельность, тем более активна данная популяция нейронов – прим пер.) в DMN ассоциировано с уменьшением этого параметра в SMN даже у здоровых людей.
Так же, как и в случае с субкортикальной модуляцией, это реципрокное взаимодействие в крайней степени выражено у пациентов с БАР в фазе депрессии и мании. У пациентов в депрессивной фазе наблюдается аномально высокая нейрональная вариабельность в DMN, что, следуя реципрокному механизму, сопровождалось снижением нейрональной вариабельности в SMN. Изменение этих параметров коррелировало с выраженностью симптоматики: чем больше была нейрональная вариабельность в DMN по сравнению с SMN, тем более выраженными были депрессивные симптомы. Интересно, что у пациентов в маниакальной фазе, наоборот, была снижена нейрональная вариабельность в DMN и, соответственно, повышена в SMN. Это также коррелировало с маниакальной симптоматикой: чем больше была вариабельность SMN по сравнению с DMN, тем более выраженные симптомы мании наблюдались у пациента. Стоит подчеркнуть, что именно отношение вариабельностей DMN/SMN коррелировало с тяжестью симптомов, но не абсолютные значения данных показателей сами по себе.
В другом исследовании Northoff et al. исследовали нейрональую вариабельность в SMN и в зрительной сети у пациентов с БАР. Авторы обнаружили, что при мании увеличение вариабельности в SMN сопровождалось уменьшением таковой в зрительной сети. Обратная картина наблюдалась при депрессии: снижение вариабельности в SMN и увеличение таковой в зрительной сети. Авторы связывали этот баланс с «внутренним» и «внешним» восприятием скорости времени. «Внутреннее» восприятие определялось активностью SMN и субкортикально-кортикальной моторной петли, в то время как «внешнее» – сенсорными областями, в том числе, зрительной сетью. Если взять степень нейрональной вариабельности в качестве показателя, отражающего восприятие времени (увеличенная вариабельность свидетельствует о большем изменении активности нейронов за единицу времени и, соответственно, об «ускорении» времени на нейрональном уровне), то можно сказать, что для пациентов с депрессией свойственно замедление «внутреннего» времени, то есть, аномальное «внутреннее замедление» (являющееся следствием уменьшения нейрональной вариабельности в SMN). Для пациентов же с манией, наоборот, характерно субъективное «ускорение» вследствие увеличенной вариабельности в SMN. Вопрос о том, каким образом эти аномалии «внутреннего времени» трансформируются в психомоторные нарушения и трансформируются ли вообще, пока остается открытым.
Данные исследования наглядно демонстрируют, что нейрональная активность в моторной коре и SMN зависит не только от субкортикальных влияний ядра шва и черной субстанции, но и от активности других, немоторных сетей коры. Самым ярким доказательством этого является реципрокное взаимодействие между DMN и SMN, а также модуляция SMN сенсорными сетями. Тем не менее, механизмы, посредством которых подобная реципрокная кортико-кортикальная модуляция ассоциирована с психомоторными функциями, еще только предстоит исследовать.
Модуляция локально-региональной активности моторной коры общей корковой активностью
Традиционно мы измеряем нейрональную активность локально, то есть, в какой-то конкретной области (амплитуда сигнала от области), либо в какой-то конкретной сети (синхронизация между собой различных отделов сети, оцениваемая при помощи rsFC). Однако в дополнение к этим локальным методам существуют также методы, позволяющие оценить активность мозга в общем. Одним из таких методов является определение rsFC между целыми нейронными сетями, например, между DMN и SMN. Кроме того, если взять вместе все межрегиональные и межсетевые связи, можно получить так называемый «глобальный сигнал», оцениваемый при помощи фМРТ. Глобальный сигнал вычисляется как среднее от всех rsFC по всему головному мозгу и отражает степень, в которой синхронизированы между собой различные области и сети, то есть глобальную синхронизацию мозга. Исследования демонстрируют, что степень, в которой та или иная область или сеть синхронизирована с остальными частями мозга, различается у разных отделов. Например, нейрональная активность в SMN более синхронизирована с общей активностью мозга (и, соответственно, демонстрирует больший глобальный сигнал на фМРТ), чем DMN, чьи отделы, по-видимому, работают более обособленно, то есть десинхронизированно (что выражается более низким глобальным сигналом на фМРТ).
В настоящее время имеются сообщения об изменениях глобального сигнала при различных психических расстройствах. Yang et al. продемонстрировали, что у пациентов с шизофренией значимо выше глобальный сигнал по сравнению с пациентами с БАР и здоровыми испытуемыми. Это означает, что общая синхронизация активности между отдельными зонами/сетями мозга аномально высокая у пациентов с шизофренией (однако в другом исследовании Argyelan et al., наоборот, продемонстрировали снижение глобальной синхронизации у пациентов с шизофренией). Более того, было обнаружено, что при шизофрении глобальная активность становится менее синхронизированной с активностью отделов/сетей низкого порядка, особенно сенсорных зон. Напротив, глобальная активность становится более синхронизированной с активностью зон высокого порядка. Wang et al. дополнили эти данные, показав, что синхронизация отдельных сетей с общей активностью мозга является динамическим, изменяющимся во времени, процессом: сначала глобальная активность синхронизируется с сенсорными сетями, затем с DMN, а после уже и с остальными сетями. Эта последовательность синхронизации глобальной активности мозга с отдельными зонами/сетями, по-видимому, нарушается при шизофрении.
В другом исследовании Zhang et al. изучали глобальный сигнал у биполярных пациентов в маниакальной, депрессивной и в эутимной фазах. Было обнаружено, что у пациентов с БАР в депрессивной фазе усилена синхронизация глобальной активности мозга с гиппокампом, что может быть ассоциировано с более частым «вызовом из памяти» автобиографических воспоминаний. Кроме того, оказалось, что у пациентов в мании глобальная активность сильно синхронизирована с активностью моторной коры, что отражает характерную для данных пациентов большую психомоторную активность. Последняя находка особенно подчеркивает, что синхронизация глобальной активности является настоящим психомоторным механизмом: изменения в моторной коре вызывают не связанные с субкортикально-кортикальной моторной петлей изменения общей активности мозга.
У данного обзора имеются следующие важные ограничения. Во-первых, обозреваемые работы сильно отличаются друг от друга по диагностическим группам, подходам к определению психомоторных нарушений и нейровизуализационным техникам. Во-вторых, разнообразие подходов и методик вкупе с небольшим количеством исследований не дали нам возможность провести полноценный ALE-мета-анализ (activation likelihood estimation – подход к мета-анализу нейровизуализационных исследований, заключающийся в определении вероятности того, что выявляемые в однородных экспериментах зоны высокой активности коры действительно совпадают – прим. пер.). В-третьих, в силу разнообразия терминологии, относящейся к концепции «психомоторного», мы могли пропустить часть важных исследований. Исходя из перечисленных выше причин, мы считаем необходимым проведение дальнейших трансдиагностических лонгитудинальных нейровизуализационных исследований психомоторных нарушений с использованием стандартизированных методов.
В данном обзоре были показаны различные нейрональные механизмы, лежащие в основе психомоторных симптомов при различных психических заболеваниях. Это является своего рода продолжением классического взгляда на психомоторные синдромы, актуальным в настоящее время, так как было продемонстрировано, что нейрональные и биохимические механизмы возникновения психомоторных нарушений не ограничиваются дофаминовой субкортикально-кортикальной моторной петлей. Мы выявили три трансдиагностических нейробиологических механизма, лежащих в основе психомоторных функций. 1) модуляция серотонином и ядром шва дофаминергической субкортикально-кортикальной моторной петли; 2) реципрокный баланс между сетью пассивного режима работы мозга, сенсомоторной и сенсорными сетями; 3) локальная синхронизация сенсомоторной сети c глобальной активностью головного мозга.
Эти механизмы в основе своей довольно похожи. Во-первых, все три механизма зависит не от каких-то абсолютных значений, а от относительных показателей, то есть, от баланса между какими-либо параметрами. Мы столкнулись с тремя видами нейронного баланса, относящимися к различным уровням проявления психомоторных функций: 1) баланс между rsFC ядра шва и черной субстанции; 2) баланс между активностью сети пассивного режима работы мозга и соматосенсорной сети; 3) баланс между глобальной активностью мозга и локальной активностью соматосенсорной сети. Кроме того, мы выявили различные виды биохимического баланса, например, между дофамином и серотонином, формирующие психомоторные функции посредством модуляции субкортикально-кортикальных и кортико-кортикальных нейронных балансов.
Во-вторых, разобранные нами данные свидетельствуют о дименсиональной и транснозологической природе психомоторных механизмов, имеющих место не только при различных заболеваниях (например, при шизофрении, БАР или депрессии), но и в норме. Соответственно, психомоторные механизмы являются иллюстрацией преимуществ применения дименсионального транснозологического синдромного подхода (как это сделано, в частности, авторами RDoc)
В-третьих, данные примеры демонстрируют континуум здоровых и патологических психомоторных состояний, в которых задействован один и тот же механизм, но проявляющийся в различной степени. В рамках данного континуума норма соответствует средним значениям, в то время как патологические состояния расположены на противоположных концах, что, таким образом, формирует кривую в форме перевернутой буквы U (см. Рис.2)
Рис.2. Перевернутые U-образные кривые континуума различных нейробиологических механизмов психомоторной активности. А. Континуум различных видов баланса между функциональной связностью ядра шва (RN) и черной субстанции (SN) с таламусом (Thal) и его связи с сенсомоторной сетью (SMN), определяющей ее активность. B. Континуум различных видов баланса между активностью SMN и сетью пассивного режима работы мозга (DMN), определяющего уровень активности SMN. C. Континуум различных видов баланса между глобальной активностью мозга (GA) и уровнем активности SMN.
В-четвертых, первернутая U-образная кривая демонстрирует, что именно средние показатели различных видов нейробиологического баланса являются наиболее оптимальными. В отличие от них, крайние формы смещения баланса являются дисфункциональными, приводящими к нарушению психомоторных функций.
В-пятых, с клинической точки зрения психомоторные нарушения могут быть охарактеризованы специфическим набором симптомов, то есть, сочетанием определенных двигательных, аффективных и когнитивных нарушений. Например, психомоторное возбуждение может сопровождаться эмоциональными нарушениями (например, повышенными аффектом) и когнитивными нарушениями (в частности, нарушениями внимания). В то же время психомоторное торможение часто сопровождается негативными эмоциями и усилением внимания на самом себе. Подобные конструкции симптомов позволяют предположить наличие связей между субкортикально-кортикальной моторной системой и немоторными, аффективными и когнитивными, нейронными системами, что подтверждается, например, существованием реципрокных взаимодействий между SMN, DMN и сенсорными сетями.
Наконец, все перечисленные механизмы имеют потенциальное диагностическое и терапевтическое значение. Они могут применяться для ранней диагностики пограничных состояний и манифеста психических заболеваний, служить потенциальными биомаркерами терапевтического ответа и быть мишенями неинвазивных методов стимуляции головного мозга (транскраниальной магнитной стимуляции и др.) Несмотря на то, что при помощи этих методов может проводиться стимуляция разных отделов мозга, все они тем или иным образом повлияют на активность субкортикально-кортикальной моторной петли и уменьшать выраженность психомоторных нарушений. Как и в случае с Римом, все дороги, в конечном счете, ведут к моторной коре.
Автор перевода: Кибитов А.А.
Источник: Northoff, G., Hirjak, D., Wolf, R.C. et al. All roads lead to the motor cortex: psychomotor mechanisms and their biochemical modulation in psychiatric disorders. Mol Psychiatry (2020). https://doi.org/10.1038/s41380-020-0814-5
Клещевой энцефалит
Данное заболевание имеет вирусную этиологию и предшествует повышению температуры, развитию головной боли и недомогания, ослабеванию мышечной ткани, на фоне чего возникает ригидность. При запущенной форме клещевого энцефалита симптоматика усугубляется: у пациента часто проявляются галлюцинации и навязчивые идеи, он становится агрессивным и возбужденным, его тревожат судороги.
Коревой
При развитии коревого энцефалита поражение больших полушарий проявляется резким повышением температуры, спутанностью сознания, в результате чего пациент не может ориентироваться во времени и пространстве, зрительными галлюцинациями и нервной возбудимостью. Характерными симптомами коревого энцефалита являются также судороги, развивающиеся во всех частях тела, парез и слабость мышц конечностей: присутствует риск возникновения коматозного состояния.
Летаргический
Если поражение коры мозга обусловлено развитием летаргического энцефалита, у пациента повышается температура, ухудшается речь, возникает кататония – патологическое состояние, при котором тело застывает в одном положении на несколько часов. Для летаргического энцефалита характерны и такие симптомы, как апатическое состояние, недомогание, сонливость, обусловленная нарушением режима дня.
Важно Обзор и применение параподиумов для реабилитации неврологических больных
Строение коры головного мозга
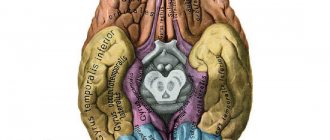
Описываемый отдел – самый крупный, он составляет до 80% всей массы представленного органа. Кора головного мозга представляет собой слой серого вещества, располагающийся на поверхности и периферии больших полушарий. Она отвечает за высшую нервную деятельность и большинство психических процессов. Весь отдел классифицируется на 5 видов долей. Каждая из них имеет собственные функции.
Лобные доли
Передняя часть коры отделена от теменной и височной двумя бороздами (центральной и латеральной). Лобные доли головного мозга состоят из нескольких основных извилин:
- предцентральная;
- верхняя лобная;
- средняя лобная;
- нижняя лобная.
Последняя извилина делится мелкими ветвями латеральной борозды на 3 части:
- оперкулярную;
- треугольную;
- орбитальную.
За что отвечают доли мозга в лобной зоне:
- мотивация;
- тормозные рефлексы;
- целенаправленное (осознанное) поведение;
- планирование;
- исполнение поставленных задач;
- способность писать;
- речь;
- точность движений.
Височные доли
Боковые зоны обладают самыми выраженными и четкими границами. Строение головного мозга в представленных долях отделяет их латеральной и двумя длинными височными бороздами (верхней и нижней). Некоторые участки «изрезаны» короткими поперечными ветвями. Височные доли мозга делятся на 3 основные извилины:
- нижняя;
- средняя;
- верхняя.
Функции этого участка коры:
- слух;
- зрительная память;
- восприятие речи;
- формирование эмоций;
- вербальная память;
- сознание;
- восприятие и сочинение музыки;
- обработка зрительной информации.
Теменные доли
Представленная область располагается вверху мозга, ближе к его задней части. Она отделена от других участков центральной и теменно-затылочной бороздой. Главной задачей рассматриваемой части коры является анализ окружающего пространства. Теменные доли головного мозга делятся на несколько областей, каждая из которых обладает специфической функциональностью:
- Первичная соматосенсорная кора.
Отвечает за восприятие и анализ тактильных ощущений. Она помогает правильно распознавать предметы, извлекать из памяти знания об их структуре, массе и форме. - Заднебоковые отделы теменных долей.
Формируют визуально-пространственные соотношения и объединяют их с другими характеристиками объектов. Данная зона обеспечивает корректное восприятие положения и траекторий перемещения своего тела и конечностей, окружающих предметов. - Среднетеменная доля в доминантном полушарии.
Функции – математические способности, письмо, понимание разницы между левой и правой стороной, узнавание пальцев. - Субдоминантная теменная доля.
Регулирует осознание собственного тела и его частей, взаимосвязь объектов в пространстве, способность объединять кусочки в целые композиции, выполнение простых механических задач (одевание, расчесывание и других).
Затылочные доли
Это образование коры локализуется у основания черепа, отделено латеральными бороздами. Задачи, которые выполняют затылочные доли мозга, и их функции, определены нервными пучками в их строении. В описываемой области сосредоточены структуры, отвечающие за зрение. Строение коры головного мозга затылочных долей включает 2 участка:
- зона визуальных ассоциаций;
- первичная зрительная кора.
Базовые функции:
- регуляция движений глаз;
- получение визуальной информации, ее обработка;
- анализ изображений;
- способность отличать предметы друг от друга визуально;
- объективное восприятие реальности.
Островковая доля
Описываемая часть коры расположена в глубине латеральной борозды. Пока это самая неизученная область в плане выполняемых функций. Предположительно островковая доля головного мозга отвечает за:
- формирование сознания;
- возникновение эмоций;
- поддержку гомеостаза;
- обработку любой сенсорной информации и ее объединение, анализ (визуальной, тактильной, аудио);
- контроль моторики;
- эмпатию;
- речь;
- восприятие тепла, холода, насыщения, полноты мочевого пузыря;
- ощущение одышки;
- равновесие;
- рефлекторные реакции (смех, плач);
- обучение движениям;
- социальные эмоции (отвращение, нормы поведения и другие);
- оргазм.
Строение
Морфологическое строение больших полушарий в пределах головного мозга предполагает наличие коркового слоя, состоящего из серого вещества, и подкорковых отделов, образованных белым веществом. Внутри массива белого вещества находятся локальные участки серого – ядра. Белое вещество служит основой для нервных волокон:
- Ассоциативных. Объединяют разные функциональные участки в рамках одной гемисферы.
- Комиссуральных. Соединяют участки разных гемисфер, чаще симметрично расположенных.
- Проводящих. Связывают отделы мозга – головного и спинного, благодаря чему образуется единая сеть ЦНС.
Строение коры, покрывающей большие полушария, предполагает наличие борозд (углублений относительно поверхности) и извилин (возвышенностей относительно поверхности). В анатомии площадь коры, покрывающей большие полушария, увеличивается за счет присутствия борозд. Продолжительность и глубина борозд – индивидуальные характеристики человека.
Корковый слой, образованный из серого вещества, содержит центры управления, которые отвечают за высшие психические функции, что взаимосвязано с такими процессами, как познавательная и мыслительная деятельность. Кора, покрывающая большие полушария в пределах головного мозга, координирует функции организма, формируя адаптивные реакции на внешние воздействия. Корковый слой образован 6-ю видами клеток.
Цитоархитектоника (расположение клеток в ткани) коры предполагает наличие молекулярной, зернистой (наружной, внутренней), пирамидной (наружной, внутренней), полиформной пластинок. В пределах головного мозга различают поверхности полушарий – верхнелатеральную (охватывает боковые верхние участки), медиальную (находится в срединной области), базальную (прилегает к основанию черепной коробки).
Верхнелатеральная поверхность обладает выпуклой формой, прилежит к костным структурам, образующим свод черепа. Плоская медиальная поверхность располагается напротив аналогичной поверхности второй гемисферы. Каждая гемисфера состоит из 5 долей. Лобная доля коры, покрывающей большие полушария – это самый большой отдел конечного мозга. Центральная борозда, анатомически поперечно пересекающая головной мозг – граница между лобной и теменной долями в рамках больших полушарий.
Сильвиева, известная так же как латеральная, борозда проходит в пределах больших полушарий перпендикулярно центральной, отделяя височную долю конечного мозга от двух сегментов – лобного и теменного. Теменно-затылочная борозда – граница теменного сегмента в задней стороне. Островковая доля в пределах гемисфер головного мозга находится в глубине сильвиевой борозды. Островковая доля просматривается, если приподнять участки лобной и височной долей, которые ее прикрывают.
Важно 3 признака развития синдрома зудека после перелома
Сильвиева, известная так же как боковая, борозда, которая отделяет участок височной доли от мозговых структур лобной и теменной – одна из самых крупных. Она образуется на 14-й неделе внутриутробного развития. Мелкие борозды, разделяющие доли коры большого мозга на извилины, формируются на 24-38 неделе, образуя своеобразный рельеф поверхности полушарий, индивидуальный для каждого человека.
Борозды и извилины 3-го порядка продолжают развиваться после рождения, наиболее интенсивно на 1-м году жизни. В лобной доле коры в пределах больших полушарий параллельно центральной пролегает прецентральная борозда, отделяющая, ограничивающая прецентральную извилину – центр, где происходит инициация и регуляция сознательной двигательной активности, направляемой волевым усилием.
Прецентральную извилину перпендикулярно пересекают лобные борозды (верхняя, нижняя). Они разделяют на сегменты лобные извилины. На заднем участке лобной извилины, расположенной снизу, находится центр Брока, который отвечает за речевую функцию. В теменной доле параллельно центральной пролегает постцентральная борозда.
Расположенная между указанными бороздами постцентральная извилина является центром чувствительности, который отвечает за реакции организма на болевые, тактильные, температурные воздействия. Височные борозды (верхняя, нижняя) пролегают в височной доле, располагаются параллельно сильвиевой борозде.
Височные борозды разделяют на сегменты височные извилины, где в верхней части располагается центр, отвечающий за функцию слуха, и центр Вернике, ответственный за речевую функцию. Шпорная борозда, которая обеспечивает зрительную функцию, пролегает в затылочной доле, где граничит с теменно-затылочной бороздой.
Литература
- Collins A., Koechlin E. Reasoning, learning, and creativity: frontal lobe function and human decision-making //PLoS biology. – 2012. – Т. 10. – №. 3. – С. e1001293.
- Chayer C., Freedman M. Frontal lobe functions //Current neurology and neuroscience reports. – 2001. – Т. 1. – №. 6. – С. 547-552.
- Kayser A. S. et al. Dopamine, corticostriatal connectivity, and intertemporal choice //Journal of Neuroscience. – 2012. – Т. 32. – №. 27. – С. 9402-9409.
- Panagiotaropoulos T. I. et al. Neuronal discharges and gamma oscillations explicitly reflect visual consciousness in the lateral prefrontal cortex //Neuron. – 2012. – Т. 74. – №. 5. – С. 924-935.
- Zelikowsky M. et al. Prefrontal microcircuit underlies contextual learning after hippocampal loss //Proceedings of the National Academy of Sciences. – 2013. – Т. 110. – №. 24. – С. 9938-9943.
- Flinker A. et al. Redefining the role of Broca’s area in speech //Proceedings of the National Academy of Sciences. – 2015. – Т. 112. – №. 9. – С. 2871-2875.
Внутри черепа есть мост
Схема строения мозга показывает, что еще одна его составляющая, варолиев мост, находится ниже ножек и образует выдающийся выступ (из-за направленных в мозжечок поперечных волокон). В передней зоне моста присутствуют преимущественно проводящие пути. А именно: путь ядер слухового нерва, вышеупомянутый пирамидальный (двигательный, корково-мышечный), общий чувствительный (от спинного мозга к зрительному бугру, медиальная петля) и др.
Строение мозга человека включает в себя такой элемент, как мозжечок. Он находится над продолговатым мозгом в задней черепной ямке. Покрыт сверху затылочными долями коры самого головного мозга. Мозжечок обладает двумя полушариями и центральной частью – червем. У новорожденного этот орган весит 20 граммов, к пяти месяцам масса увеличивается в 3 раза, к девяти месяцам – в 4 раза.
Окончательно формирование этого органа завершается к 15 годам. Поверхность полушарий представляет собой серое вещество – кору, под которой скрывается белое вещество. Орган имеет три пары ножек, связывающих его с другими отделами: нижняя пара – с продолговатым мозгом, средняя – с варолиевым мостом, верхняя — со средним мозгом.
Белое вещество
Оно представлено в виде многочисленных волокон. Их разделяют на три группы:
- Проекционные. Эта категория представлена пучками эфферентных и афферентных волокон. Посредством них существуют связи между проекционными центрами и базальными, стволовыми и спинномозговыми ядрами.
- Ассоциативные. Эти волокна обеспечивают соединение корковых участков в границах одного полушария. Они разделены на короткие и длинные.
- Комиссуральные. Эти элементы связывают корковые зоны противоположных полушарий. Комиссуральными образованиями считаются: мозолистое тело, задняя и передняя спайка и спайка свода.
Префронтальная кора
Крупный отдел коры больших полушарий, который представлен в виде передних отделов лобных долей. С ее помощью осуществляется контроль, управление, фокусировка любых действий, которые совершает человек. Этот отдел позволяет нам правильно распределять своё время. Известный психиатр Т. Голтьери охарактеризовал этот участок в качестве инструмента, с помощью которого люди ставят цели, разрабатывают планы. Он был уверен, что правильно работающая и хорошо развитая префронтальная кора – важнейший фактор эффективности личности.
К основным функциям префронтальной коры также принято относить:
- Концентрацию внимания, сосредоточение на получении только необходимой человеку информации, игнорирование сторонних мыслей и чувств.
- Способность «перезагружать» сознание, направляя его в нужное мыслительное русло.
- Настойчивость в процессе выполнения определенных задач, стремление к получению намеченного результата, несмотря на возникающие обстоятельства.
- Анализ складывающейся в настоящий момент ситуации.
- Критическое мышление, позволяющее создать комплекс действий для поиска проверенных и достоверных данных (проверка полученной информации перед ее использованием).
- Планирование, выработка определенных мер и действий для достижения поставленных целей.
- Прогнозирование событий.
Отдельно отмечается способность этого отдела управлять эмоциями человека. Здесь процессы, протекающие в лимбической системе, воспринимаются и переводятся в конкретные эмоции и чувства (радость, любовь, желание, горе, ненависть и т.д.).