Functions of the striopallidal system, limbic system and cerebral cortex
No. 1.1. Functions of the globus pallidus. Afferent and efferent connections
The globus pallidus (globus pallidus s. pallidum) has predominantly large type I Golgi neurons. Connections between the globus pallidus and the thalamus, putamen, caudate nucleus, midbrain, hypothalamus, somatosensory system, etc. indicate its participation in the organization of simple and complex forms of behavior.
Stimulation of the globus pallidus using implanted electrodes causes contraction of the muscles of the limbs, activation or inhibition of γ-motoneurons of the spinal cord. In patients with hyperkinesis, irritation of different parts of the globus pallidus (depending on the location and frequency of irritation) increased or decreased hyperkinesis.
Stimulation of the globus pallidus, unlike stimulation of the caudate nucleus, does not cause inhibition, but provokes an orienting reaction, movements of the limbs, feeding behavior (sniffing, chewing, swallowing, etc.).
Damage to the globus pallidus causes in people hypomimia, mask-like appearance of the face, tremor of the head and limbs (and this tremor disappears at rest, during sleep and intensifies with movements), and monotony of speech. When the globus pallidus is damaged, myoclonus is observed - rapid twitching of the muscles of certain groups or individual muscles of the arms, back, and face.
In the first hours after damage to the globus pallidus in an acute experiment on animals, motor activity sharply decreased, movements were characterized by incoordination, the presence of incomplete movements was noted, and a drooping posture occurred when sitting. Having started moving, the animal could not stop for a long time. In a person with dysfunction of the globus pallidus, the onset of movements is difficult, auxiliary and reactive movements disappear when standing up, friendly movements of the arms when walking are disrupted, and a symptom of propulsion appears: long-term preparation for movement, then rapid movement and stopping. Such cycles are repeated many times in patients.
Most of the afferent signals coming to the basal ganglia enter the striatum of the basal ganglia. These signals come almost exclusively from three sources: from all areas of the cerebral cortex, from the intraplate nuclei of the thalamus and the substantia nigra of the basal ganglia. Efferent fibers from the striatum go to the globus pallidus of the basal ganglia and the substantia nigra, from which the dopaminergic pathway to the striatum and pathways to the thalamus begin. The most important of all efferent tracts originates from the internal part of the globus pallidus, ending in the thalamus and the roof of the midbrain.
No. 1.2. Functions of the caudate nucleus and putamen. Afferent and efferent connections
The caudate nucleus (nucleus caudatus) and the shell (putamen) are evolutionarily later formations than the globus pallidus and functionally have an inhibitory effect on it.
The caudate nucleus and putamen have a similar histological structure. Their neurons belong to type II Golgi cells, i.e. they have short dendrites and a thin axon; their size is up to 20 microns. These neurons are 20 times more numerous than type I Golgi neurons, which have an extensive network of dendrites and are about 50 microns in size.
The functions of any brain formation are determined primarily by their connections, of which the basal ganglia have quite a lot. These connections have a clear focus and functional outline.
The caudate nucleus and putamen receive descending connections primarily from the extrapyramidal cortex through the subcallosal fasciculus. Other areas of the cerebral cortex also send large numbers of axons to the caudate nucleus and putamen.
The main part of the axons of the caudate nucleus and putamen goes to the globus pallidus, from here to the thalamus, and only from it to the sensory fields. Consequently, there is a vicious circle of connections between these formations. The caudate nucleus and putamen also have functional connections with structures lying outside this circle: with the substantia nigra, red nucleus, Lewis body, vestibule nuclei, cerebellum, γ -cells of the spinal cord.
The abundance and nature of the connections between the caudate nucleus and the putamen indicate their participation in integrative processes, the organization and regulation of movements, and the regulation of the work of vegetative organs.
Irritation of field 8 of the cerebral cortex causes excitation of neurons in the caudate nucleus, and field 6 causes excitation of neurons in the caudate nucleus and putamen. A single stimulation of the sensorimotor area of the cerebral cortex can cause excitation or inhibition of the activity of neurons in the caudate nucleus. These reactions occur within 10-20 ms, which indicates direct and indirect connections between the cerebral cortex and the caudate nucleus.
The medial nuclei of the thalamus have direct connections with the caudate nucleus, as evidenced by the reaction of its neurons, which occurs 2-4 ms after stimulation of the thalamus.
The reaction of neurons in the caudate nucleus is caused by skin irritations, light and sound stimuli. —
In the interactions between the caudate nucleus and the globus pallidus, inhibitory influences prevail. If the caudate nucleus is irritated, most of the neurons of the globus pallidus are inhibited, and a smaller part is excited. If the caudate nucleus is damaged, the animal develops motor hyperactivity.
The interaction of the substantia nigra and the caudate nucleus is based on direct and feedback connections between them. It has been established that stimulation of the caudate nucleus increases the activity of neurons in the substantia nigra. Stimulation of the substantia nigra leads to an increase, and destruction leads to a decrease in the amount of dopamine in the caudate nucleus. It has been established that dopamine is synthesized in the cells of the substantia nigra and then transported at a speed of 0.8 mm/h to the synapses of neurons in the caudate nucleus. In the caudate nucleus, up to 10 mcg of dopamine accumulates in 1 g of nervous tissue, which is 6 times more than in other parts of the forebrain, the globus pallidus, and 19 times more than in the cerebellum. Thanks to dopamine, a disinhibiting mechanism of interaction between the caudate nucleus and the globus pallidus appears. With a lack of dopamine in the caudate nucleus (for example, with dysfunction of the substantia nigra), the globus pallidus is disinhibited, activating the spinal-stem systems, which leads to motor disorders in the form of muscle rigidity.
Corticostriatal connections are topically localized. Thus, the anterior regions of the brain are connected to the head of the caudate nucleus. Pathology that occurs in one of the interconnected areas of the cortex - the caudate nucleus, is functionally compensated by the preserved structure. Corticostriatal connections are topically localized. Thus, the anterior regions of the brain are connected to the head of the caudate nucleus. Pathology that occurs in one of the interconnected areas of the cortex - the caudate nucleus - is functionally compensated by the preserved structure.
The caudate nucleus and globus pallidus take part in such integrative processes as conditioned reflex activity and motor activity. This is detected by stimulation of the caudate nucleus, putamen and globus pallidus, destruction and by recording electrical activity.
Direct stimulation of some zones of the caudate nucleus causes the head to turn in the direction opposite to the irritated hemisphere, the animal begins to move in a circle, i.e., a so-called circulatory reaction occurs.
Irritation of other areas of the caudate nucleus and putamen causes the cessation of all types of human or animal activity: orientation, emotional, motor, food. At the same time, slow-wave activity is observed in the cerebral cortex.
In humans, stimulation of the caudate nucleus during a neurosurgical operation disrupts speech contact with the patient: if the patient said something, he becomes silent, and after the irritation stops he does not remember that he was addressed. In cases of brain injury with irritation of the head of the caudate nucleus, patients experience retro-, antero- or retroanterograde amnesia.
In animals such as monkeys, stimulation of the caudate nucleus at different stages of the implementation of a conditioned reflex leads to inhibition of the execution of this reflex. For example, if the caudate nucleus of a monkey is stimulated through implanted electrodes before giving a conditioned signal, then the monkey does not react to the signal, as if it had not heard it; irritation of the nucleus after the monkey goes to the feeder at the signal or already begins to take food from the feeder leads to the animal stopping; after the irritation stops, the monkey, without completing the conditioned reaction, returns to its place, i.e. it “forgets” that it was irritating signal (retrograde amnesia).
Stimulation of the caudate nucleus can completely prevent the perception of painful, visual, auditory and other types of stimulation. Irritation of the ventral region of the caudate nucleus reduces, and the dorsal region increases salivation.
When the caudate nucleus is stimulated, the latent periods of reflexes lengthen and the alteration of conditioned reflexes is disrupted. The development of conditioned reflexes against the background of stimulation of the caudate nucleus becomes impossible. Apparently, this is explained by the fact that stimulation of the caudate nucleus causes inhibition of the activity of the cerebral cortex.
A number of subcortical structures also receive the inhibitory influence of the caudate nucleus. Thus, stimulation of the caudate nuclei caused fusiform activity in the thalamus optic, globus pallidus, subthalamic body, substantia nigra, etc.
Thus, specific for irritation of the caudate nucleus is primarily inhibition of the activity of the cerebral cortex, subcortical formations, inhibition of unconditioned and conditioned reflex behavior.
At the same time, when the caudate nucleus is irritated, some types of isolated movements may appear. Apparently, the caudate nucleus has, along with inhibitory and excitatory structures.
Switching off the caudate nucleus is accompanied by the development of hyperkinesis such as involuntary facial reactions, tremor, athetosis, torsion spasm, chorea (twitching of the limbs and torso, as in an uncoordinated dance), motor hyperactivity in the form of aimless moving from place to place.
In case of damage to the caudate nucleus, significant disorders of higher nervous activity, difficulty in orientation in space, memory impairment, and slowed growth of the body are observed. After bilateral damage to the caudate nucleus, conditioned reflexes disappear for a long period of time, the development of new reflexes becomes difficult, general behavior is characterized by stagnation, inertia, and difficulty switching. In monkeys after unilateral damage to the caudate nucleus, the conditioned reaction was restored after 30-50 days, the latent periods of reflexes were lengthened, and intersignal reactions appeared. Bilateral damage led to complete inhibition of conditioned reflexes. Apparently, bilateral damage depletes symmetrical compensatory mechanisms.
When affecting the caudate nucleus, in addition to disorders of higher nervous activity, movement disorders are noted. Many authors note that in different animals, with bilateral damage to the striatum, an uncontrollable desire to move forward appears, while with unilateral damage, manege movements occur.
Despite the great functional similarity between the caudate nucleus and the putamen, there are a number of functions specific to the latter. evolutionarily, the shell appears earlier than the caudate nucleus (its rudiments are already present in fish).
The shell is characterized by participation in the organization of eating behavior: food search, food orientation, food capture and food possession; a number of trophic disorders of the skin and internal organs (for example, hepatolenticular degeneration) occur when the function of the shell is impaired. Irritation of the shell leads to changes in breathing and salivation.
As mentioned earlier, irritation of the caudate nucleus inhibits the conditioned reflex at all stages of its implementation. At the same time, irritation of the caudate nucleus prevents the extinction of the conditioned reflex, i.e., the development of inhibition; the animal ceases to perceive the new environment. Considering that stimulation of the caudate nucleus leads to inhibition of the conditioned reflex, one would expect that destruction of the caudate nucleus would cause facilitation of conditioned reflex activity. But it turned out that the destruction of the caudate nucleus also leads to inhibition of conditioned reflex activity. Apparently, the function of the caudate nucleus is not simply inhibitory, but lies in the correlation and integration of RAM processes. This is also confirmed by the fact that information from different sensory systems converges on the neurons of the caudate nucleus, since most of these neurons are polysensory.
No. 1.3. Functional organization of the nigro-striopallidal system
The striopallidal system is divided according to its functional significance and morphological features into striatum and pallidum (Table 3).
Table 3. Functional and morphological differences between the striatum and pallidum
No other system is as susceptible to the external modeling influence of hormones and neurotransmitters as the extrapyramidal system, since functionally it is part of the structure of the limbic system. That is why the energetic mood of a person’s movements depends on emotions, and when the subcortical ganglia are damaged, affective disorders (violent laughter and crying) are revealed. Functions of the extrapyramidal system.
1. Regulation of muscle tone in conjunction with other structures.
2. Regulation of tempo, rhythm and plasticity of any voluntary motor act.
3. Providing a motor component in the regulation of unconditioned reflexes (sexual, defensive, start reflex, etc.).
4. Ensuring the sequence of the motor act.
5. Providing the motor component of the emotional sphere.
6. Regulation of highly specialized human movements that have reached the level of automatism.
No. 1.4. Symptoms of damage to the nigro-striopallidal system
Pallidum involvement: pallidal syndrome
The symptom complex of damage to the globus pallidus and substantia nigra is called parkinsonism, akinetic-rigid syndrome, amyostatic syndrome, hypertensive-hypokinetic syndrome. It is associated with a functional deficiency of dopamine, with a change in the influence of the pallidonigral system on the reticular formation and a violation of impulses in the cortical-subcortical-stem neural circles. The reticular formation is the stem “controller-regulator” of the flow of ascending and descending impulses. When its connections with the substantia nigra are disrupted, it does not prevent the passage of excess tonic signals to the muscle, as a result of which muscle rigidity develops, supported by a continuous flow of afferent impulses to the striopallidal system. A vicious circle arises: the affected pallidal system sends uncontrolled tonic signals that increase muscle tone and increase the flow of reverse afferentation impulses.
The main symptoms of damage to the globus pallidus are:
• changes in muscle tone according to the plastic type, the “gear wheel” phenomenon (the resistance experienced during the study of tone increases towards the end of the movement), plastic rigidity;
• pose of a wax doll, mannequin - a phenomenon of catalepsy (when moving from a resting position to a state of movement, patients often freeze in an uncomfortable position);
• the “petitioner” pose is the characteristic appearance of patients: the torso is slightly bent, the head is tilted forward (Fig. 4.2), the arms are bent and brought to the body, the gaze is directed forward, motionless;
• oligokinesia - poverty and lack of expressiveness of movements; hands are motionless when walking, there is no normal physiological synkinesis;
• bradykinesia - slowness of movements; patients are inactive, inert, constrained;
• parkinsonian treading on the spot - the onset of a motor act is difficult, so at the beginning of each movement the patient makes several repeated movements, for example, rocking or walking in place;
• inertial movement forward - propulsion, to the side - lateropulsion, backward - retropulsion; a patient who is out of balance cannot restore it automatically;
• bradyllia - monotonous, slow, quiet speech;
• bradypsychia - slowing down the pace of thinking;
• micrography - small, unclear handwriting;
• akairia - “viscosity” in communication, stickiness;
• “paradoxical kinesia” - patients who sit in a chair all day, at the time of affective outbursts and emotional stress, can run up the stairs, jump, dance;
• parkinsonian rest tremor: most often localized in the fingers - the phenomenon of “rolling pills”, “counting coins”, head tremor - the phenomenon of “yes-yes, no-no, no-yes”; trembling is observed at rest and decreases with voluntary movements;
• positive postural reflexes: the Westphal foot phenomenon (with passive dorsiflexion of the foot, tonic tension of the foot extensors occurs - it freezes in the dorsiflexion position) and the leg phenomenon (in a patient lying on his stomach with his legs bent at a right angle at the knees, with further passive flexion of the lower leg, it freezes in the flexion position);
• sleep-wake rhythm disturbance;
• autonomic disorders (greasy face, peeling skin, hypersalivation).
Damage to the striatum: striatal syndrome
When the striatal system is damaged, a dystonic-hyperkinetic syndrome occurs, caused by a deficiency of the inhibitory influence of the striatum on the underlying motor centers, as a result of which muscle hypotonia and excessive involuntary movements (hyperkinesis) develop.
Hyperkinesis - automatic, excessive movements that involve individual parts of the body and limbs. They arise involuntarily, disappear during sleep and intensify with voluntary movements and excitement.
Athetosis - slow, worm-like, fanciful movements in the distal parts of the limbs (in the hands and feet) [Fig. 4.3]. Athetosis in the facial muscles is accompanied by protrusion of the lips, distortion of the mouth, grimacing, and clicking of the tongue. Athetosis is usually associated with damage to large cells of the striatal system. Its characteristic feature is the formation of transient contractures (spasmus mobilis), which give the hand and fingers a peculiar position.
Ballism, hemiballism - large, sweeping, “throwing” movements of the limbs. Most often, ballism affects the muscles of the arms, causing a movement similar to the flapping of a bird's wing. Violent movements during hemiballismus are made with great force and are difficult to stop. The occurrence of hemiballismus is associated with damage to the body of Lewis, located under the optic thalamus.
Chorea is rapid contractions of various muscle groups of the face, trunk and limbs. Hyperkinesis is irregular, uncoordinated, and extends to large muscles of the distal and proximal sections. May resemble voluntary movements, as synergists are involved in the process. Frowning of the eyebrows and forehead, protruding tongue, and jerky, erratic movements of the limbs are noted. Hyperkinesis covering half the body is called hemichorea. Chorea occurs when the neostriatum is damaged and is observed with subcortical degenerations, rheumatic brain damage, and Huntington's disease.
In some cases, choreic hyperkinesis is combined with athetosis (choreoathetosis). Choreoathetosis can be observed in patients both constantly and in the form of attacks - paroxysmal choreoathetosis. Several variants of the familial form of paroxysmal choreoathetosis have been described.
Dystonia is forced muscle contractions, leading to “twisting” or hyperextension of a part of the body. There is spastic torticollis (local dystonia of the neck muscles), in which the head is turned to the side and tilted towards the shoulder (Fig. 4.4). Involuntary head tilts forward or backward are also possible. At the beginning of the disease, muscle tension is transient, but over time it becomes permanent, as a result of which the head is always in an unnatural position. Spasmodic torticollis often occurs in the initial stages of torsion dystonia as a local manifestation. Torsion dystonia is a generalized variant of hyperkinesis. Movements of the body are rotational, corkscrew-like in nature, accompanied by hyperlordosis, scoliosis, and pretentious poses. Torsion dystonia can stop with various compensatory techniques, for example, by wrapping your arms around the neck, increasing rotation of the shoulder, etc. Dystonia of other muscle groups is also distinguished.
• Writer's cramp (graphospasm) is a convulsive contraction of the fingers that occurs during writing.
• Occupational cramps are spasms of muscles involved in certain occupational movements. Observed in violinists, pianists, guitarists, typists, etc.
• Facial hemispasm - periodically repeated spasms of the muscles of half the face innervated by the facial nerve. Convulsions are accompanied by the appearance of wrinkles on the forehead, the corner of the mouth is pulled outward and upward, and the platysma is tense.
• Meij's facial paraspasm is recurrent symmetrical spasms of the facial muscles. Paraspasm often occurs during a conversation or smile.
Blepharospasm is a convulsive contraction of the orbicularis oculi muscle. Clinically, blepharospasm is manifested by frequent blinking and occurs paroxysmally.
Hiccups are clonic spasms of the diaphragm. It manifests itself with fast, loud inhalation movements and is caused by a pathological process in the membranes or in the substance of the brain, intoxication.
Myoclonus is a rapid hyperkinesis that looks like a wince. There are local myoclonus (for example, limbs) and generalized. One should also distinguish non-epileptic myoclonus (hyperkinesis) from epileptic myoclonus.
Tics are rapid clonic twitches of a limited muscle group, usually of a stereotypical nature, imitating voluntary movements. They are most often localized in the facial muscles and are manifested by rapid wrinkling of the forehead, raising of the eyebrows, blinking, and sticking out the tongue. The tic of the neck muscles is accompanied by turning the head to the side and nodding. In children, a tic often develops as a manifestation of neurosis as a result of a pathological conditioned reflex, as an imitation of persons suffering from hyperkinesis. Facial tics can occur with trigeminal neuralgia. Tics can involve the muscles of the limbs, trunk, and diaphragm. A special place is occupied by a generalized impulsive tic - de la Tourette's syndrome, in which impulsive jumping, squatting, grimacing, vocal phenomena in the form of grunting, screaming, shouting swear words (coprolalia) are observed.
Trembling (tremor) is a stereotypical clonic rhythmic hyperkinesis, mainly observed in the hands and feet; trembling of the torso and head may also be noted. Trembling is an external manifestation of often invisible muscle contraction as a result of damage to segmental and suprasegmental motor structures, primarily the striopallidal system and the cerebellum. The amplitude of the jitter and its frequency, the duration of individual phases may vary depending on the mechanism of occurrence. There is a large-scale, “rubral” tremor that occurs when the red nucleus is damaged and affects the opposite half of the body. The tremor may increase or decrease with movement. Intention tremor sharply intensifies or occurs when performing purposeful movements, in particular when performing coordination tests - finger-nose and heel-knee. Static tremor is observed at rest, does not increase with movements (on the contrary, it may decrease), and is characteristic of parkinsonism.
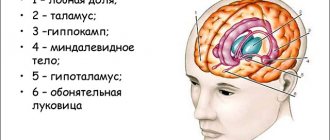
No. 2. Composition and functions of the limbic system
The limbic system is a functional union of brain structures involved in the organization of emotional and motivational behavior.
What you need to know about the basal ganglia
The large hemispheres of the human brain on the outside are a cortex formed by gray matter, and on the inside by a subcortex of white matter.
The basal ganglia (ganglia, nodes), which are also called central or subcortical, are concentrations of gray matter in the white matter of the subcortex. The basal ganglia are located at the base of the brain, which explains their name, outside the thalamus (optic thalamus). These are paired formations that are symmetrically represented in both hemispheres of the brain. With the help of nerve processes, they interact bilaterally with various areas of the central nervous system.
The main role of the subcortical nodes is to organize motor function and various aspects of higher nervous activity. Pathologies that arise in their structure affect the functioning of other parts of the central nervous system, causing problems with speech, coordination of movements, memory, and reflexes.
Features of the structure of the basal ganglia
The basal ganglia are located in the frontal and partially temporal lobes of the telencephalon. These are clusters of neuron bodies that form groups of gray matter. The white matter surrounding them is represented by processes of nerve cells and forms layers that separate the individual basal ganglia and other brain structural and functional elements.
The basal nodes include:
- striatum;
- fence;
- amygdala.
In anatomical sections, the striatum appears as alternating layers of gray and white matter. It consists of caudate and lenticular nuclei. The first is located anterior to the visual thalamus. As the caudate nucleus becomes thinner, it becomes the amygdala. The lenticular nucleus is located lateral to the thalamus optic and the caudate nucleus. It is connected to them by thin bridges of neurons.
The fence is a narrow strip of neurons. It is located between the lenticular nucleus and the insular cortex. It is separated from these structures by thin layers of white matter. The amygdala is shaped like the amygdala and is located in the temporal lobes of the telencephalon. It consists of several independent elements.
This classification is based on the structural features and location of the ganglia on an anatomical section of the brain. There is also a functional classification, according to which scientists classify only the striatum and some ganglia of the diencephalon and mesencephalon as the basal ganglia. These structures collectively provide human motor functions and individual aspects of behavior responsible for motivation.