Speed of propagation of nerve impulses
In 1830, one of the greatest physiologists of the 19th century, Johann Muller, stated that the speed of propagation of PD was impossible to measure. In his opinion, since PD is an electrical impulse, it should be carried out at a speed approximately equal to the speed of light (3–1010 cm/s); Given the small size of biological objects, even with the best instruments of the time it was impossible to measure such a speed.
15 years later, one of Müller’s students, Hermann von Helmholtz, using a simple and elegant experiment that can be easily reproduced in a student laboratory workshop (Fig. 6–8), measured the speed of propagation of impulses in the nerve of a frog. In his experiments, Helmholtz irritated the nerve in two areas spaced 3 cm apart and measured the time from the moment the stimulus was given to the maximum muscle contraction. Let's assume that when the distal (located closer to the muscles) area is irritated, this time decreases by 1 ms. Then the pulse propagation speed V
equal to
| V | = | d | / | t | = | 3 cm | / | 1 ms | = | · | 103 | cm/s |
| Rice. 6.8. An experimental setup similar to the one with which Helmholtz measured the speed of propagation of impulses in the frog nerve. The stimulating electrodes were first applied to point St1 and then to point St2. A lever was connected to the muscle, the pointed end of which drew a curve on a sooty sheet of paper, which was quickly moved in the longitudinal direction. |
This value turned out to be seven orders of magnitude less than the speed of propagation of electric current in a copper conductor or in an electrolyte solution. From this, Helmholtz made the absolutely correct conclusion that the conduction of a nerve impulse is a more complex process than the simple longitudinal propagation of current in a nerve fiber.
The speed of impulse propagation in different axons varies from 120 m/s (in some large fibers) to several centimeters per second (in very thin axons). These differences between the conduction speed in different fibers are illustrated in Table. 6–1 and fig. 6–9.
The speed of impulse propagation largely depends on how quickly a section of the membrane located at a certain distance from the site of stimulus delivery is depolarized by local currents to a threshold level. The higher the fiber length constant, the further these currents can propagate, the faster the depolarization of the membrane in front of the excitation site occurs and, therefore, the higher the speed of impulse propagation. The effect of constant length on this speed can be demonstrated by placing the axon in oil or air. In this case, only a thin film of saline solution remains on the surface of the axon, and the length constant decreases due to an increase in the external longitudinal resistance [in equation (6–2) – r
0]. Under these conditions, the speed of excitation will be lower than when the axon is immersed in a saline solution.
Table 6-1. Classification of frog nerve fibers according to their diameter and speed of excitation (Erlanger, Gasser, 1937)
| Fiber group | Diameter, microns | Speed, m/s |
| A α | 18,5 | |
| β | 14,0 | |
| γ | 11,0 | |
| B | — | 4,2 |
| C | 2,5 | 0,4 –0,5 |
| Rice. 6.9. The speed of propagation of excitation in various groups of frog nerve fibers. A. Experimental setup for stimulating a bundle of nerve fibers and recording the resulting potentials. B. Composite action potential recorded using extracellular electrodes and representing the sum of potentials in all excited fibers of the bundle. Group α fibers have the largest diameter and are characterized by the highest conduction speed. In contrast, group γ fibers have both the diameter and conduction velocity the lowest (see Table 6-1). Stimulation was carried out until the start of recording. |
In the process of evolution, living organisms have developed two ways to increase the constant length of the axon and thereby the speed of impulse propagation. One of them (a typical example would be giant axons of squids, arthropods, annelids, and teleost fishes) is an increase in the diameter of the axon, i.e., a decrease in the internal longitudinal resistance [in equation (6–2) – r
i] This issue is discussed in more detail in Appendix 6-2. Giant axons have evolved in some animal species to provide rapid synchronous activation of motor reflexes, such as mantle movements in squid and the withdrawal or avoidance reflex in some arthropods (crayfish, cockroaches) and annelids (e.g., earthworms).
The work of neurotransmitters
When they transmit nerve impulses, there are several options for what will happen to them:
- They will diffuse.
- Will undergo chemical breakdown.
- They will return back to their bubbles (this is called recapture).
At the end of the 20th century, an amazing discovery was made. Scientists have learned that drugs that affect neurotransmitters (as well as their release and reuptake) can radically change a person's mental state. For example, a number of antidepressants like Prozac block the reuptake of serotonin. There are some reasons to believe that a deficiency in the brain neurotransmitter dopamine is to blame for Parkinson's disease.
Now researchers who study the borderline states of the human psyche are trying to figure out how all this affects the human mind. Well, for now we do not have an answer to such a fundamental question: what causes a neuron to create an action potential? For now, the mechanism for “launching” this cell is a secret to us. Particularly interesting from the point of view of this riddle is the work of neurons in the main brain.
In short, they can work with thousands of neurotransmitters sent by their neighbors. The details regarding the processing and integration of this type of impulses are almost unknown to us. Although many research groups are working on this. At the moment, we have learned that all received impulses are integrated, and the neuron makes a decision whether it is necessary to maintain the action potential and transmit them further. The functioning of the human brain is based on this fundamental process. Well, then it is not surprising that we do not know the answer to this riddle.
Saltatory conduction
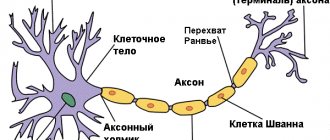
The second way to increase the speed of nerve impulses, realized only in vertebrates, is to isolate sections of the axon using the myelin sheath.
In this case, the length constant of the corresponding sections increases significantly, and thereby the conduction of current in the longitudinal direction is significantly facilitated. As the animal develops, myelin is deposited around peripheral and central axons by glial cells located near these axons. As a result, a dense multilayer shell of cell membranes is formed around the fibers.
Cells that synthesize myelin include Schwann cells
(in the area of peripheral nerves) and
oligodendrocytes
(Fig. 6–10) (in the central nervous system).
Transverse sections of the myelin sheath show periodically repeating gaps of 12 nm, resulting from the layering of glial cell membranes. With the formation of each new layer, the lateral resistance of the shell increases. Since there are many layers in this shell, its capacity is much lower than that of a single membrane. The multilayer myelin sheath is periodically interrupted (the so-called nodes of Ranvier},
and in these small areas the excitable axon membrane is in contact with the extracellular environment. Between the nodes of Ranvier, the myelin sheath is closely adjacent to the axon membrane, practically displacing the extracellular environment. In addition, sections of the axon membrane between the nodes of Ranvier , apparently, do not contain sodium channels.
| Rice. 6.10. Interception of Ranvier. Shown is a short "bare" section of an axon located between two myelinated regions. It is this area that is excited during saltatory conduction. In Fig. 4–12 shows an electron micrograph showing a multilayer myelin sheath formed by the membranes of glial cells. (Bunge et al., 1961.) |
Due to the insulating properties of the myelin sheath, the axon length constant increases sharply: the presence of this sheath has the same effect as an increase in r
m [Equation (6–2)]. Due to the high resistance of the myelin sheath, local currents flowing ahead of the excitation wave exit the axon almost exclusively in the region of the nodes of Ranvier. In addition, since the capacitance of the thick myelin sheath is small, only a very small current is consumed to recharge this capacitance in the areas between nodes.
Due to these features, the AP arising in any interception electrotonically depolarizes only the membrane located in the area of the next interception, and therefore impulses in such axons do not propagate along their entire length, as in unmyelinated nerve fibers (for example, in the squid axon). They arise only in small areas of the membrane—the nodes of Ranvier. All this causes saltatory
(spasmodic)
conduction,
in which impulses propagate intermittently from interception to interception (Fig. 6–11). Speed of spread. In this case, the PD increases sharply, since the electrotonic conduction of local currents between interceptions occurs very quickly. Thus, in vertebrates, Nature solved the problem of the rapid propagation of nerve impulses without resorting to the creation of such bulky structures as giant axons.
How does everything work in the brain?
Let's talk about what transmission sequence of nerve impulses works in the most important parts of our central nervous system. Here, neurons are separated from their neighbors by small gaps called synapses. The action potential cannot pass through them, so it looks for another way to get to the next nerve cell. At the end of each process there are small sacs called presynaptic vesicles. Each of them contains special compounds - neurotransmitters. When an action potential arrives at them, molecules are released from the sacs. They cross the synapse and attach to special molecular receptors that are located on the membrane. In this case, the equilibrium is disturbed and, probably, a new action potential appears. This is not yet known for certain; neurophysiologists are still studying the issue to this day.
Nerve impulse transmission speed
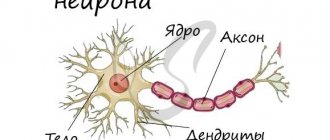
Nerve impulse, propagation of excitation (bioelectric impulse) along nerve fibers in response to irritation of neurons.
In the second half of the 19th century, in the work of G. Helmholtz and E. Hering on the frog nerve, it was shown that the bioelectric signal (current, or action potential), unlike the electric current in a conventional conductor, propagates along the nerve fiber with a finite speed (3- 120 m/sec).
The possibility of propagation of nerve impulses along nerve fibers is determined by their structure, which is reminiscent of the structure of an electric cable, where the role of a conductor is played by axons, and the role of an insulator is the axon’s myelin sheath, which is a Schwann cell membrane wound around the axon in several layers.
The main component of the myelin sheath is the lipoprotein myelin, which has dielectric properties. The speed of propagation of nerve impulses depends both on the diameter of the nerve fibers (the thicker the fiber, the higher the speed), and on the degree of their electrical insulation, since myelin-coated fibers, other things being equal, conduct nerve impulses faster. The myelin sheath does not cover the fiber continuously along its entire length, but forms a kind of insulating ceramic “couplings” tightly strung on the axon, like on the rod of an electrical cable.
Between adjacent “couplings” of myelin, only small electrically non-insulated areas remain, through which ionic current can easily flow from the axon into the external environment and back, irritating the membrane and causing the generation of an action potential exclusively in non-insulated areas of the axon, called nodes of Ranvier. The nerve impulse spreads along the myelinated nerve fiber in jumps - from one node of Ranvier to the next, which significantly increases the speed of propagation of excitation from cell to cell.
The speed of propagation of a nerve impulse along thick myelinated fibers (10-20 microns in diameter) in humans reaches 70-120 m/sec, and along the thinnest unmyelinated nerve fibers it is two orders of magnitude lower (less than 2 m/sec).
The ability to produce nerve impulses is one of the fundamental properties of neurons.
Nerve impulses provide rapid transmission of similar signals (action potentials) along axons over long distances and therefore are the most important means of exchanging information both between nerve cells and between nerve and other types of cells. Information about the strength of stimulation of a nerve cell is encoded and transmitted to other cells by changing the frequency of nerve impulses.
The repetition rate can vary from a few to hundreds of nerve impulses per second. The frequency code assumes a complex periodicity of nerve impulses, including their grouping into “packs” with different numbers and patterns of signals. The complex spatial and temporal summation of nerve impulses forms the basis of the rhythmic electrical activity of the brain, recorded using an electroencephalogram.
The speed of propagation of nerve impulses can vary: less than 1 meter per second in very thin axons and about 100 meters per second in thick axons (for example, in axons innervating muscles).
An electrical impulse propagating along the axon, reaching the axon endings on another nerve cell, suddenly disappears. Charles Sherrington, who laid the foundations of the so-called synaptology, called the points of contact between the endings of an axon and another nerve cell “synapses.”
In order to “cross” a synapse, the nerve impulse must be regenerated on the other side of the synapse. Even 15 years ago, some physiologists believed that the transmission of impulses through a synapse is a phenomenon mainly (if not entirely) of an electrical order. Now, however, there is ample evidence that during such transmission there is a release of special substances that cause impulse regeneration. The first convincing evidence that a transmitter substance operates in a synapse was obtained more than 40 years ago by G. Dale and O. Löwy.
As is known, the human central nervous system (including, of course, not only the brain, but also the spinal cord) consists of approximately 10 billion (1010) nerve cells. Almost all nerve cells, with rare exceptions, receive information directly in the form of impulses (see figure below) from several nerve cells at once (often hundreds of them) and transmit it to an equally large number of cells.
About the action potential
Speaking about the transformation of a nerve impulse and its propagation, it should be noted that it could amount to measly millimeters per second. Then signals from the hand to the brain would take minutes, which is clearly not good. This is where the previously discussed myelin sheath plays its role in enhancing the action potential. And all its “passes” are placed in such a way that they only have a positive effect on the speed of signal transmission. So, when an impulse reaches the end of the main part of one axon body, it is transmitted either to the next cell or (if we talk about the brain) to numerous branches of neurons. In the latter cases, a slightly different principle works.
Excitation and inhibition of a nerve cell
Excitation and inhibition of a nerve cell are carried out by nerve fibers that form synapses on its surface.
Top (1) motor neuron at rest. Impulses arriving along one excitatory fiber (2) are not yet able to cause a discharge of the motor neuron. A discharge occurs only when impulses also arrive along the second excitatory fiber (3) (threshold state of the neuron). If the neuron also receives impulses along the inhibitory fiber, then it returns to the subthreshold state (4).
Bottom (b) - impulses arrive only along the inhibitory fiber. Electrical impulses propagating along excitatory and inhibitory nerve fibers do not differ from each other. Their opposite action is explained by the release of different chemical transmitters in the synaptic endings.
In a given nerve cell, depending on its excitation threshold, a discharge of impulses may occur when only a few fibers coming to it are irritated; in other cases, the discharge of impulses does not occur even when many such fibers are irritated.
It has long been known that various factors can increase or decrease the threshold of excitation of a nerve cell.
Moreover, about 60 years ago it was suggested that some fibers should inhibit the discharge of impulses in the cell to which they approach, rather than excite it. This assumption was subsequently confirmed, and at present the mechanism of inhibition is clarified. This article is devoted to two equivalent processes—inhibition and its antipode—excitation of a nerve cell.
An interesting aspect of the activity
Speaking about the conduction of nerve impulses by cells, it is difficult not to talk about one interesting point. So, when they are at rest, then, let's say, the sodium-potassium pump is engaged in moving ions in such a way as to achieve the effect of fresh water inside and salty outside. Due to the resulting imbalance, potential differences across the membrane can be observed up to 70 millivolts. For comparison, this is 5% of regular AA batteries. But as soon as the state of the cell changes, the resulting equilibrium is disrupted, and the ions begin to change places. This happens when the path of a nerve impulse passes through it. Due to the active action of ions, this action is also called an action potential. When it reaches a certain point, reverse processes begin and the cell reaches a state of rest.
Some theoretical features
In the article, "nerve impulse" and "action potential" were used as synonyms. In theory this is true, although in some cases it is necessary to take into account some features. So, if you go into detail, the action potential is only part of the nerve impulse. With a detailed examination of scientific books, you can find out that this is only the name for a change in the charge of the membrane from positive to negative, and vice versa. Whereas a nerve impulse is understood as a complex structural-electrochemical process. It spreads across the neuron membrane as a traveling wave of change. An action potential is just the electrical component of a nerve impulse. It characterizes the changes that occur with the charge of a local area of the membrane.