The medulla oblongata is localized in the posterior part of the brain and connects it with the spinal part of the central nervous system. Formed from the structures of the rhombencephalon and is part of the brainstem. Controls the primary processes of the body - breathing and blood circulation.
Since the medulla oblongata connects the cerebrum and the spinal part of the central nervous system, it combines the functions and structure of both structures.
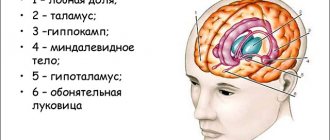
Brain in section
Structure of the medulla oblongata
The oblongata section is located at the junction of the spinal and cephalic sections of the central nervous system. Anatomically, it is located in the area of exit of the roots of the 1st cervical spinal nerve, and is limited from above by the bridge. In appearance it resembles an onion. Its height in an adult is approximately 2-3 cm.
Despite its relatively small size, it performs many functions without which the individual could not exist.
Its structures consist of white and gray matter, which determines its dual functions - it combines the characteristics of the spinal and telencephalon with minor differences: its internal part consists of compactly located clusters of gray matter, and the external part is made of white matter. From the back of the oblong section, 2 cords stretch to the dorsal section, which unite them.
As you know, the cerebral hemispheres control opposite sides of the body: the left side responds to the right side and vice versa. This feature of the central nervous system is achieved due to the characteristic structure of the medulla oblongata: it implements a conductor function, which consists of transmitting excitation impulses from its own nuclei to the nervous tissue of other parts, but not in a direct way, but crosswise.
The size and structure of the medulla oblongata change with age. For example, unlike adults, in infants this part of the central nervous system is larger relative to other parts. It acquires its final appearance by the age of 7 years.
Location boundaries
External structure
The structure of the medulla oblongata is divided into 2 parts: internal and external. Let us take a closer look at the appearance of this department and the features of its structure from an anatomical point of view.
The medulla oblongata is an inverted cone: its wide end is at the top, the narrow end is at the bottom. In the central part of the front side there is an anterior median fissure. It is a continuation of the central sulcus of the dorsal part of the central nervous system.
On the sides of it there are pyramids that pass into the facial cables of the spinal cord, consisting of a cluster of nerve cells. On the posterior side of the medulla oblongata, the posterior middle sulcus is visible, which passes into the dorsal region.
Detailed structure
Next to it are the caudal cords, which are connected to the grooves of the spinal part of the central nervous system. These cords are the lower boundary line of the oblong section, and the basal boundary, passing in the area of the Varoliev bridge, has become its upper boundary.
Internal structure
The internal structure of the medulla oblongata is more effectively viewed in a cross section made at the level of the olives. If you look at it from the front side, the anatomy looks like this: tracts of intentional motor impulses extend from paired elevations (pyramids) on the outer side into the depths of the structure.
Next to them are olives, covering the basal ganglia, the roots of the hypoglossal nerve going to the lingual muscles, axons and nerve cells grouped into nuclei. Long fibers pass from each nucleus to the cerebellum, connecting them to each other. They form the olivocerebellar tract.
Near the olive tree there are also:
- Fibers of the glossopharyngeal nerve;
- Vagus nerve fibers;
- Accessory nerve fibers.
The efferent conduction tracts lie on the lateral (side) side of the medulla oblongata:
- Rubrospinal;
- Tectospinal;
- Reticulospinal;
- Vestibulospinal.
At the level of the pyramids, most of the axons (up to 80%) that form the efferent pathways intersect with each other. The rest also form a decussation and move down to the other side, to the spinal part of the central nervous system.
In the posterior part of the medulla oblongata, 4 sensory tracts stretch from the spinal cord to the upper parts of the central nervous system:
- Thin;
- Wedge-shaped;
- Spinothalamic;
- Spinocerebellar.
They are a natural continuation of similar structures of the spinal cord.
How to work with this lesson?
I have finally completed this medulla oblongata anatomy guide and can now focus on making a guide to the guide. In fact, there are several points that will greatly facilitate your study of neuroanatomy in general, and the structure of the medulla oblongata in particular:
- First learn the anatomy of the spinal cord, read my spinal cord guides carefully;
- Correlate the external structure of the medulla oblongata with the external structure of the spinal cord;
- Be sure to draw the pathways that connect the spinal cord and medulla oblongata;
- Remember not only the names and location, but, above all, the functions of each nucleus and each pathway;
- Use Latin, it makes memorization much easier;
- Use video tutorials from masters of normal anatomy such as Professor Izranov and Professor Kafarov.
Explanation of point 3
As you can see, I drew the pathways in this lesson very schematically, deliberately omitting very important features - first of all, this concerns the nuclei of the spinal cord. For example, if we were to completely draw a segment of the spinothalamic tract, we would have to draw this in detail:
- The impulse is met by the first neuron, which is located in the spinal ganglion (1);
- Then, along the process of this neuron it enters its own nucleus, which lies in the dorsal horn (2);
- Next, along the neuron process from its own nucleus, the impulse enters the medulla oblongata and follows to the thalamus, where the third nucleus is located.
As you can see, this would create unnecessary clutter, since it would be necessary to add anatomical structures that are not directly related to the medulla oblongata. I will most likely do a separate lesson on pathways, but to do that we need to study all parts of the CNS.
The lines that connect the medulla oblongata and the spinal cord in this lesson very roughly and schematically show the pathways. I drew them so that, while studying the medulla oblongata, you would constantly correlate each new element with the spinal cord. This will help you a lot in the future.
Main nuclei of the medulla oblongata
The gray matter of the medulla oblongata processes data arriving in the central nervous system from all receptors of the human body. It is a cluster of neuron structures (bodies and partly processes), compactly located in the thickness of the white matter.
Highlight:
- Nuclei of 9-12 pairs of cranial nerves;
- Circulatory and respiratory centers;
- Olive kernel;
- Reticular formation.
In the medulla oblongata there are 5 pairs of cranial nerve nuclei. They are concentrated in the caudal part of the cerebrum below the bottom of the 4th ventricle:
- Vestibulocochlear nerve (VIII pair). Consists of the cochlear and vestibular parts. Responsible for conducting auditory impulses, as well as signals originating from the vestibular part of the inner ear.
- Glossopharyngeal nerves (IX pair). Includes sensory, motor and secretory fibers. Motor axons implement motor function - they control the activity of the muscles of the pharyngeal canal and oral cavity. With the help of sensory fibers, the nucleus receives information received from the food-gustatory sensory system of the back of the tongue. Parasympathetic fibers are responsible for the secretory function in the oral cavity: they control salivary processes.
- Vagus nerve (X pair). Consists of 3 types of nuclei: vegetative, sensory and motor. The autonomic nucleus functions as a connecting link between the cerebrum and the organs of the head, neck, chest and abdominal cavity. Incoming and descending fibers extend from it in different directions. The sensitive core receives information from the lungs, heart and other important systems of the body. The motor nucleus controls contraction of the oral muscles during swallowing. The reciprocal nucleus is also located here, the axons of which perform a protective function - they activate the processes of coughing, sneezing, and vomiting.
- Accessory nerve (XI pair). It is represented by 2 nuclei: one lies in the medulla oblongata at the bottom of the rhomboid fossa, the other in the anterior horn at the level of 1 segment of the spine. Fibers from it enter the foramen magnum of the skull, unite with the cranial roots, forming the trunk of the accessory nerve. They are then brought out from the cranial cavity through the jugular foramen. The accessory nerve performs a controlling function - it is responsible for the motility of the sternocleidomastoid and trapezius muscles.
- Hypoglossal nerve (XII pair). Controls the motor function of the tongue, as it is associated with its styloglossus, mentalis, rectus and transverse muscles. Unconditioned reflexes depend on its work: swallowing, chewing, sucking. The nuclei of the hypoglossal nerve control the motor skills of the tongue during chewing, speaking, and other mouth movements. Information from the hypoglossal nerve is sent to the sensorimotor region of the cerebral cortex through the sphenoid and tender nucleus.
The hemodynamic center is located in the lower caudal part of the oblongata. Its fibers are closely connected with the nuclei of the V pair of nerves. Scientists suggest that it is in the hemodynamic center that excitatory activating signals of sympathetic fibers to the cardiovascular system arise.
Core kernels
Sometimes the hemodynamic center is suppressed by shock, trauma, stroke, poisoning, and metabolic disorders.
Olive kernel. It is a serrated plate, the functions of which are determined by its structure - it consists of gray matter. Curved in the shape of a horseshoe. Interacts with the dentate nuclei of the cerebellum, that is, they are the main structures of balance.
The olivary kernel is the best developed of all mammals in humans, since humans, more than anyone else in the animal world, are in an upright position and need a good vestibular apparatus. In addition, the olive nuclei help to navigate in space: they compare the volume of sound waves penetrating the right and left ear. Analyzing them, a person turns to face the source of the sound.
In the lower part of the medulla oblongata there is the so-called “blue spot” - the zone of the reticular formation. The fibers of neurons of this structure secrete a specific hormone that affects the degree of excitability of nerve cells.
The “blue spot” is responsible for feelings of anxiety and internal tension, and also ensures the department’s connection with the sensory organs, the spinal cord and other departments, serving as a regulator of nervous activity.
3.4. BRAIN AND PONTUS
3.4.1. Structure of the medulla oblongata and pons
The medulla oblongata and the pons (pons) are a part of the brain that is a direct continuation of the spinal cord. The gray matter of the spinal cord passes into the gray matter of the medulla oblongata and retains the features of a segmental structure. However, the main part of the gray matter is distributed throughout the entire volume of the hindbrain in the form of isolated nuclei,
separated by white matter.
The nuclei of the last four pairs of cranial nerves, which carry out afferent and efferent innervation of the head and internal organs, are symmetrically located in the medulla oblongata. The motor nucleus of the hypoglossal nerve is located in the lowermost part of the rhomboid fossa.
(XII pair), innervating the muscles of the tongue.
The motor nucleus of the accessory nerve
(XI pair) is
| Rice. 3.21 Location of cranial nerve nuclei in the brainstem 1 - accessory nerve, 2 - hypoglossal, 3 - vestibulocochlear, 4 - abducens, 5 - trochlear, 6 - oculomotor, 7 - trigeminal, 8 - facial, 9 - glossopharyngeal, 10 - vagus nerve. |
continuation of the posterolateral section of the ventral horn of the spinal cord and is associated with the innervation of the neck muscles (Fig. 3.21).
vagus nerve nuclei lies quite superficially
(X pair), which is mixed, i.e., simultaneously involved in autonomic, motor and sensory innervation.
Accordingly, the function of the vagus nerve belongs to three nuclei. The autonomic parasympathetic nucleus of the vagus nerve
produces parasympathetic fibers going to the larynx, esophagus, heart, stomach, small intestine, and digestive glands (see section 5.3).
Sensory fibers of the vagus nerve from many internal organs in the medulla oblongata form the solitary tract (solitary tract) and end in the sensory nucleus of the solitary tract.
The third nucleus of the vagus nerve - somatic, motor - lies deeper in the lateral part of the medulla oblongata and is called
double.
The neurons of this nucleus reflexively regulate the sequence of contractions of the muscles of the pharynx and larynx during breathing.
Glossopharyngeal nerve
(IX pair), like the vagus, contains motor, sensory and autonomic fibers. Motor fibers are involved in the innervation of the muscles of the oral cavity and pharynx. Sensitive fibers from the taste buds of the posterior third of the tongue and the mucous membrane of the pharynx enter together with the fibers of the vagus nerve into the solitary tract. The processes of neurons that form the autonomic nucleus of the glossopharyngeal nerve go to the parasympathetic ganglia, which are associated with the innervation of the secretory apparatus of the salivary glands.
At the border of the medulla oblongata and the pons are the nuclei of the vestibulocochlear,
or
auditory
nerve (VIII pair). This nerve consists of two parts:
cochlear and pre-door. The fibers of the cochlear part, or the auditory nerve itself, are axons of the cells of the spiral ganglion lying in the cochlea. These fibers end in the cochlear nuclei
(Fig. 3.21).
Fibers of the vestibule
are formed by the axons of the cells of the vestibule node, localized at the bottom of the internal auditory canal.
These fibers end in the three vestibular nuclei of the bridge: medial
(Schwalbe's nucleus),
lateral
(Deiters' nucleus) and
superior
(Bechterew's nucleus). The processes of second-order neurons from these nuclei are directed to the cerebellar tent nucleus. From the lateral vestibular nucleus, which can be classified as a suprasegmental formation, the vestibular cord begins, leading to the spinal cord.
The nucleus of the facial nerve is located in the lateral part of the reticular formation of the pons
(VII pair). This nerve is mixed. Afferent sensory fibers of the facial nerve transmit information from the taste buds of the anterior part of the tongue. Autonomic fibers participate in the innervation of the submandibular and sublingual salivary glands. The efferent fibers of the facial nerve have terminal branches in the facial muscles.
Abducens nerve nucleus
(VI pair) is laid superficially in the posterior half of the bridge. The motor fibers of this nerve innervate the muscles that move the eyeball, and the sensory fibers are connected to the proprioceptors of these muscles.
And finally, the last pair of cranial nerves belonging to the hindbrain is the trigeminal nerve
(V pair).
This nerve is mixed and has motor and sensory nuclei. The motor nucleus of the trigeminal nerve
is located in the upper part of the pontine tegmentum.
The neurons of this nucleus are involved in the innervation of the masticatory muscles, the velum palatine muscle and the tensor tympani muscle. The long sensory nucleus of the spinal tract of the trigeminal nerve,
, contains cells to which afferent axons from receptors on the skin of the face, nasal mucosa, teeth, and periosteum of the skull bones approach (see Fig. 3.21).
The gray matter of the medulla oblongata contains the nuclei of the sensory pathways of the posterior columns of the spinal cord. These are thin and wedge-shaped nuclei (nuclei of bundles of the same name),
which are pathways that carry information from tactile and proprioceptors, signaling the position of the body in space.
In addition, located within the hindbrain is the posterior nucleus of the trapezius body
(superior olivary nucleus), which is included in the ascending pathway, which transmits acoustic information (see section 4.5.3). Along the lateral surface of the medulla oblongata there is a continuation of the lateral columns of the spinal cord.
The central part of the medulla oblongata is occupied by the reticular,
or reticular,
a formation
(substance), which is a collection of nerve cells, reminiscent of the principle of organization of the nerve networks of coelenterates.
Reticular neurons are distributed diffusely and grouped in nuclei, among which the largest can be distinguished: giant cell reticular, small cell reticular, lateral reticular nucleus at the level of the medulla oblongata, caudal, ventricular and oral reticular nuclei at the level of the bridge. According to the cytoarchitectonic features in the reticular formation of the medulla oblongata, a medial part
with many large neurons and a lateral part, which includes smaller elements, are distinguished. According to its topography, the reticular formation of the brain stem is a structure located along the path of all input and output systems of the brain.
The dendrites and soma of reticular neurons are approached by collaterals from the axons of sensory ascending pathways localized in the central and lateral parts of the brain stem. On the other hand, collaterals of descending motor pathways coming from the forebrain approach the neurons of the reticular formation. From the neurons of the giant cell and caudal reticular nucleus, the descending reticular-spinal cord begins, establishing direct connections with the motor neurons of the spinal cord. The reticular formation has connections with the nuclei of the cranial nerves, with the cerebellum, diencephalon and, through the latter, with the cerebral cortex.
Thus, the richness of connections of the reticular formation indicates that it is a suprasegmental structure adapted to regulate the activity of various parts of the central nervous system.
3.4.2. Reflexes of the medulla oblongata
The medulla oblongata is a vital part of the nervous system, where the arcs of a number of somatic and autonomic reflexes close. With the participation of the nuclei of the medulla oblongata, chain reflexes are carried out,
associated with chewing and swallowing food. For example, during the act of chewing, the coordinated activity of the masticatory muscles, tongue, cheeks, palate and floor of the mouth is ensured by the function of the motor nuclei of the trigeminal and hypoglossal nerves. When a bolus of food touches the roof of the mouth, the base of the tongue, or the back of the throat, a chain swallowing reflex occurs. From the receptors of the posterior third of the tongue, the mucous membrane of the pharynx, excitation along the sensory fibers of the glossopharyngeal nerve enters the medulla oblongata to the center of the swallowing reflex. From this center, motor commands programmed in the nervous system along the motor fibers of the hypoglossal, trigeminal, glossopharyngeal and vagus nerves go to the muscles of the oral cavity, pharynx, larynx and esophagus. As a result of the coordinated activity of these muscles, the act of swallowing occurs with simultaneous closure of the entrances to the nasopharynx and larynx.
Many autonomic reflexes
hindbrain. These include, for example, reflex regulation of the secretion of the salivary glands, which is carried out with the participation of the nuclei of the facial and glossopharyngeal nerves containing parasympathetic neurons. The autonomic, parasympathetic nucleus of the vagus nerve is included in the system of reflex regulation of breathing, cardiac activity and vascular tone.
A whole group of hindbrain reflexes is aimed at maintaining postural tone.
Depending on the source of receptor influences, tonic reflexes are divided into cervical and vestibular, or labyrinthine.
Cervical tonic reflexes
triggered by stimulation of the proprioceptors of the neck muscles.
They were first described by the Dutch physiologist R. Magnus in cats with the brain stem cut at the level of the cerebellar tent (see section 3.5.2). In their pure form, cervical reflexes can be observed when the vestibular apparatus is destroyed, which provides additional information about the position of the head in space. As the experiments of R. Magnus showed, tilting the cat’s head back in the sagittal plane causes an increase in the tone of the extensor muscles of the forelimbs and at the same time a weakening of the extensor tone of the hind limbs (Fig. 3.22). On the contrary, tilting the head down causes the opposite process, when the tone of the extensors of the forelimbs decreases, and the tone of the hind limbs increases. Impairment of balance due to turning the head to the right or left relative to the longitudinal axis causes a compensatory increase in the tone of the extensors of those limbs towards which the head is turned. This is the so-called Magnus rotation reflex.
The effector element of neck reflexes can be not only the muscles of the limbs, but also the eye muscles. Reflex influences on the eye muscles ensure compensatory adjustment of the eyeballs when the position of the head changes. Any deviation of the head causes the eyeballs to move in the opposite direction, and thus the correct visual orientation of the animal is reflexively maintained.
All neck reflexes are polysynaptic. Impulses from the proprioceptors of the cervical muscles go to the corresponding centers of the medulla oblongata, and from there descending commands arrive either to the motor neurons of the spinal cord or to the motor nucleus of the abducens nerve and produce a certain effect on postural tone and on the ocular muscles.
| Rice. 3.22 Postural reflexes of a decerebrate animal A - decerebrate rigidity; B, C, D - cervical tonic reflexes according to Magnus; see text for explanation. |
Vestibular reflexes
are inextricably linked with cervical tonic reflexes and, in natural conditions, complement them. Vestibular reflexes do not depend on the position of the head relative to the body and in their pure form can be obtained by fixing the head in relation to the body or by turning off the proprioceptors of the cervical muscles with novocaine blockade.
Vestibular reflexes, according to Magnus’ classification, are divided into static and statokinetic. Static reflexes
are associated mainly with the excitation of the receptors of the vestibule of the membranous labyrinth and ensure the maintenance of posture and balance of the body in a wide variety of its static positions in space. So, for example, if a cat with its head fixed relative to the body is rotated in space, then depending on the position of the head, a different distribution of postural tone will be observed. In the case when the angle between the horizontal plane and the plane of the animal’s oral fissure is 45°, the extensor muscles are tensed to the maximum. If the indicated angle is 135°, the lowest value of extensor tone is observed. Between the extreme positions there is a series of gradual transitions in the degree of development of extensor tone. This is an example of a tonic vestibular position reflex, which is carried out with the participation of neurons of the lateral vestibular nucleus (Deiters) and the vestibular cord going from it to the spinal cord.
Static vestibular reflexes include straightening reflexes, aimed at transitioning the animal from an unnatural position to its usual position. An illustrative example is a cat turning over in the air when falling backwards. In a very short time of falling, the animal manages to take a position normal in relation to the gravitational field and fall on all four paws at once. During the process of turning over, righting reflexes occur in a certain sequence. First, due to the vestibular rectifying reflex, the normal position of the head is restored - face down. Then the change in the position of the head excites the proprioceptors of the neck muscles and they trigger the cervical rectifying reflex, as a result of which, following the head, the torso also returns to its normal position.
Thus, under natural conditions, vestibular straightening reflexes are complemented by cervical ones, and the leading role in their implementation is played by the position of the head, where distant receptors are located.
The next group of vestibular reflexes - statokinetic -
characterized by the fact that it is aimed at maintaining posture when the speed of movement of the animal changes. These reflexes are associated with excitation of the receptors of the semicircular canals, which occurs when there is endolymph flow in the canals. Typically, the direction of the statokinetic reflex depends on which semicircular canal is irritated during acceleration. For example, when the forward movement slows down, the receptors of the sagittal canal are excited and, accordingly, reflex changes in muscle tone will tilt the torso forward, as if maintaining uniform movement.
Acceleration during rotation of the body in the horizontal plane excites the receptors of the horizontal semicircular canal and causes a reflex reaction of the oculomotor system - horizontal nystagmus.
The essence of this reaction is that at the moment of acceleration of rotation, the eyes move in the direction opposite to the direction of rotation. Then, having reached extreme deviation, the eyes quickly move back in the direction of rotation and thus another part of space enters the field of view. Nystagmus helps maintain normal visual orientation and is usually used for diagnostic purposes to check the normal functioning of the vestibular system. The plane of nystagmus coincides with the plane of acceleration, and therefore, in addition to horizontal nystagmus, you can find vertical, diagonal and circular nystagmus (see section 4.8.4).
Statokinetic vestibular reflexes also include the so-called elevator reflexes,
which manifest themselves in an increase in the tone of the extensor muscles with linear acceleration upward and in an increase in the tone of the flexors with linear acceleration downward.
3.4.3. Functions of the reticular formation of the brainstem
The reticular formation of the medulla oblongata contains centers that play an important role in the regulation of visceral functions.
This is primarily
a respiratory center
located in the medial part of the reticular formation of the medulla oblongata.
Even in the last century, it was established that damage to the caudal part of the bottom of the fourth ventricle leads to respiratory arrest (Flourance injection).
Later, N.A. Mislavsky (1885) identified two functionally different parts in the respiratory center:
inspiratory and expiratory
(or inspiratory and expiratory). The use of microstimulation methods and microelectrode recording of the activity of single neurons made it possible to clarify ideas about the localization of parts of the respiratory center.
It has been established that the inspiratory part is located more ventrally than the expiratory part. The activity of single neurons in the respiratory center can correlate with the phases of the respiratory cycle. This feature allows us to distinguish inspiratory neurons, which generate action potentials in the initial phase of inspiration, and expiratory neurons, the discharge of which is confined to the exhalation phase. The largest concentration of inspiratory neurons was found near the solitary tract. In the area of the double nucleus, inspiratory and expiratory neurons are localized. A certain mosaic pattern in the arrangement of respiratory neurons indicates that the division of the respiratory center into two parts - inhalation and exhalation - is more valid in a functional sense than in an anatomical one (see Section 9.4.1).
A distinctive feature of the neurons of the respiratory center is the ability to automatize.
Even in the absence of afferent influences, the activity of these neurons is characterized by periodicity, which is determined by the specific ionic mechanisms of their cell membrane. The periodicity of discharges of respiratory neurons may also be due to the presence of mutual inhibitory connections between inspiratory and expiratory neurons. Inhibitory connections create reciprocity of discharges of respiratory neurons, when the appearance of activity of inspiratory neurons is accompanied by inhibition of expiratory discharges and vice versa. As a result of the reciprocal interaction of respiratory neurons, a change in the phases of the respiratory cycle occurs.
At the same time, changing the phases of the respiratory cycle can be carried out reflexively due to afferent influences that modulate the respiratory rhythm. With intense inhalation and stretching of the lung tissue, impulses from the mechanoreceptors of the lungs along the afferent fibers of the vagus nerve enter the respiratory center and cause reflex inhibition of inspiratory neurons, while simultaneously stimulating expiratory ones (Hering-Breuer reflex).
Impulses from expiratory neurons along the reticular-spinal tract reach the executive motor centers of the spinal cord and stimulate the onset of exhalation.
Periodicity in the work of the respiratory center of the medulla oblongata can also be ensured due to regulatory influences from the pneumotaxic center.
This center is located in the area of the pons and also consists of two types of neurons - inspiratory and expiratory, which do not have their discharges strictly confined to the phases of the respiratory cycle. The pneumotaxic center, as a regulator, periodically inhibits the inspiratory part of the respiratory center and stimulates expiratory neurons, thus stopping inhalation and beginning exhalation.
Higher levels of the brain can also take part in regulating the functions of the respiratory center. For example, a person’s emotional reactions are associated with changes in the periodicity of the respiratory center, caused by impulses in the diencephalon and limbic cortex. It has been established that electrical stimulation of the orbital cortex can cause a complete cessation of breathing in a person. The regulatory influences of the brain provide voluntary regulation of breathing and its correction during various changes in life activity.
Thus, the regulation of respiratory function
carried out by a hierarchical system of nerve centers located on different floors of the nervous system and connected by unity of action.
Another vital center of the reticular formation of the medulla oblongata is the vasomotor center,
the localization of which was first determined by F.V. Ovsyannikov (1871). The vasomotor, or vasomotor, center occupies a large area of the medulla oblongata, extending in the dorsolateral direction from the bottom of the IV ventricle to the pyramids (see section 5.5.2). Transection of the spinal cord at the level of the cervical vertebrae causes in spinal animals a sharp decrease in vascular tone and disruption of regulatory reactions associated with changes in the lumen of veins and arteries.
Local stimulation of the rostral parts of the vasomotor center causes an increase in vascular tone, increased blood pressure and tachycardia. Stimulation of the caudal part of the vascular center, on the contrary, causes vasodilation, a drop in blood pressure and bradycardia.
Microelectrode recording of the electrical activity of single neurons of the vascular center showed the presence of nerve cells that change their background activity synchronously with fluctuations in blood pressure. Among them, we can distinguish neurons whose firing frequency increases with increasing blood pressure, neurons whose frequency of background activity, on the contrary, decreases with increasing blood pressure, and, finally, neurons whose firing frequency changes in accordance with the operating cycle of the heart. The properties of the neurons of the vascular center are less studied than those of the neurons of the respiratory center. It is not yet clear whether vasomotor neurons can be differentiated into “vasoconstrictor” and “vasodilator” neurons. The question of the presence of inhibitory interaction between neurons of the vasomotor center also remains unresolved.
The functional organization of the vasomotor center has certain specificity in terms of the projections of its descending pathways. Efferent fibers from its neurons descend into the thoracic spinal cord, but end there not on motor neurons, as in the case of the descending systems of the respiratory center, but on preganglionic neurons of the sympathetic nervous system.
Thus, vascular tone is regulated not by antagonistic descending influences, but only by the sympathetic vasoconstrictor system.
The active state of this system causes vasoconstriction, and inhibition causes vasodilation (see sections 5.4, 8.6). The only exceptions to this principle are some vessels that have double innervation - sympathetic and parasympathetic (vessels of the genital organs and salivary glands).
Reflex influences on the neurons of the vasomotor center are carried out by excitation of chemo- and mechanoreceptors localized in the vascular wall. Afferent fibers from these receptors, as part of the vagus and glossopharyngeal nerves, reach the medulla oblongata. Excitation of the mechanoreceptors of the aortic arch and carotid sinus with an increase in blood pressure causes inhibition of the activity of the vasomotor center and, as a consequence, a reflex decrease in vascular tone (Ludwig-Zion, Hering, Bainbridge reflexes). On the contrary, with increasing pressure in the vena cava system, an increase in the activity of the vascular center and a vasoconstrictor effect is observed. The tone of the vascular center can also change when the chemoreceptors of the vascular wall are excited, which occurs when the chemical composition of the blood changes.
It should be noted that the activity of the vasomotor center is combined with the function of the motor nucleus of the vagus nerve - the double nucleus, which normally reduces the heart rate. In this regard, with the vasoconstrictor effect, the heart rate simultaneously increases and, conversely, with vasodilation, a slowdown in heart rate is observed.
The reticular formation of the brainstem is characterized not only by autonomic regulatory functions, but also by participation in the descending control of the activity of the motor centers of the spinal cord.
In 1862, I.M. Sechenov established the inhibition of spinal reflexes upon irritation of the brainstem.
This was the discovery of central inhibition and at the same time the discovery of the reticulospinal system. However, the mechanism of this inhibitory process was discovered only in the 50s. of our century after the work of the American neurophysiologist G. Megun, who showed that local electrical stimulation of the giant cell nucleus of the reticular formation of the medulla oblongata causes nonspecific inhibition of flexion and extension spinal reflexes. These nonspecific supraspinal influences along the reticular-spinal tract reach the motor neurons of the spinal cord and increase the threshold and latent period of their responses to reflex influences. Further studies showed that the descending influences of the reticular formation can be realized not only due to postsynaptic inhibition of motor neurons, but also due to the occurrence of long-term inhibitory postsynaptic potentials in interneurons, as well as due to the influence of reticulospinal fibers on the terminals of afferent fibers entering the spinal cord.
Thus, the weakening of reflex activity is achieved through a direct effect on motor neurons and due to a certain weakening of sensory input into the spinal cord.
During experiments with local stimulation of the reticular formation, the presence of zones was revealed that give an effect of the opposite polarity, i.e., a facilitating effect on spinal reflexes.
For example, electrical stimulation of the lateral zones of the reticular formation of the pons lowers the threshold and shortens the latent period of spinal reflexes. When stimulating the medial nuclei of the reticular formation of the hindbrain, short-latency excitatory postsynaptic potentials (EPSPs) are recorded in the motoneurons of the cat flexor muscles.
This fact indicates the presence (in addition to diffuse nonspecific reticulospinal projections) of monosynaptic descending pathways of specific action that are involved in the descending control of spinal cord activity.
Consequently, the reticular formation, as one of the motor centers of the brain stem, can act not only as a regulator of the excitability of spinal motor neurons, but also take part in processes associated with maintaining posture and organizing purposeful movements.
Thanks to the work of G. Megun and J. Moruzzi, along with the nonspecific descending influences of the reticular formation of the trunk, its ascending, activating influences on the cerebral cortex were discovered.
If the central parts of the reticular formation of the trunk are irritated through chronically implanted electrodes, the cat, which is in a sleepy state, awakens and has an indicative reaction.
This behavioral reaction of awakening is accompanied by characteristic changes in the frequency spectrum of the electroencephalogram, a transition from regular, high-voltage oscillations of the α rhythm to low-voltage oscillations. (β-rhythm (see Section 3.11.1). This electroencephalographic reaction is called the desynchronization reaction.
It is generalized in nature and is recorded from large areas of the cerebral cortex.
In an acute experiment, transection of the brainstem at the level of the midbrain and, thus, destruction of the ascending pathways from the reticular formation of the brainstem transfers the animal into a sleep-like comatose state (sleeping brain,
according to
Bremer)
with corresponding changes in the nature of the electroencephalogram.
The above experimental facts served as the basis for the conclusion that the reticular formation is the structure responsible for the state of wakefulness, the structure that forms the ascending activating reticular system, which maintains the excitability of the diencephalon and the cerebral cortex at a certain level. According to modern concepts, the transition of the cortex to an active state is associated
| Rice. 3.23 Afferent and efferent connections of the reticular formation of the brainstem |
With
fluctuations in the number of ascending signals from the reticular formation of the trunk. The number of these signals depends on the entry of sensory impulses into the reticular formation along the collaterals of specific afferent ascending pathways. Almost to the reticular formation information comes from all sensory organs along collaterals from the spinal-reticular tract, propriospinal tracts, afferent cranial nerves, from the thalamus and hypothalamus, from the motor and sensory areas of the cortex (Fig. 3.23).
Microelectrode recording of the electrical activity of neurons of the reticular formation showed that most of them are polysensory,
i.e., they respond to stimulation of various modalities (light, sound, tactile, etc.).
Reticular neurons have large receptive fields, a long latent period, and poor response reproducibility. These properties are opposite to the properties of neurons of specific nuclei and allow reticular neurons to be classified as nonspecific.
Likewise, the ascending pathways of the reticular formation, in contrast to the classical specific sensitive projections, are called
nonspecific projections.
The ascending influences of the reticular formation are highly sensitive to the action of various pharmacological anesthetics and so-called sedatives (aminazine, serpazil, reserpine, etc.).
The use of electrophysiological and histofluorescent methods made it possible to establish the transmitter specificity of neurons in the reticular formation of the medulla oblongata, pons and midbrain. For example, the localization of monamine neurons, which begin to fluoresce after microsections of the brainstem were treated in formaldehyde vapor, was studied. It turned out that the most significant group of adrenergic
cells is located in one of the areas of the bridge called the "locus coeruleus". The axons of neurons in this region pass through the middle forebrain fasciculus and have terminal endings in various parts of the brain: the cerebellum, hypothalamus, limbic system, and cerebral cortex.
Bodies serotonergic
neurons are located predominantly in the middle region of the brain stem - as part of the dorsal and medial raphe nuclei of the medulla oblongata, midbrain and pons. The ascending axons of these neurons go to the hypothalamus, basal ganglia, limbic system, and cerebral cortex.
Currently, significant factual material has been accumulated indicating the multidirectional influence of these two monoaminergic systems on the excitability of various parts of the brain. The serotonin system induces sleep and is directly related to the regulation of its slow-wave phase. A lack of serotonin in the brain causes forced wakefulness. The catecholamine system of the locus coeruleus, on the contrary, stimulates awakening and plays a leading role in the regulation of the paradoxical phase of sleep. When the locus coeruleus is damaged and, accordingly, there is a lack of norepinephrine, animals sleep much longer than usual.
The reciprocity of monoaminergic systems is manifested not only in the processes of sleep regulation, but also in the impact on the emotional sphere of the body. For example, an increase in the concentration of cerebral norepinephrine entails an increase in the stress state of animals, which can be relieved (aborted) by the introduction of the initial product of serotonin synthesis - 5-hydroxytryptophan. Likewise, destruction of the raphe nuclei, where serotonin is synthesized, increases the aggressiveness of experimental rats.
Multidirectional influences of monoaminergic systems of the brain occur both in relation to the regulation of motor activity (the serotonin system reduces it) and in the processes of formation of complex forms of behavior (development of conditioned reflexes). This indicates the modulating function of these systems, which have extensive connections with various parts of the brain.
Other mediator systems within the reticular formation of the brainstem are represented by cholinergic neurons, as well as accompanying glycinergic cells, which most often perform the functions of inhibitory interneurons.
It should be noted that the ascending pathways of the reticular formation contain activating and deactivating groups. According to J. Moruzzi, irritation of certain areas of the reticular formation of the hindbrain can cause deep sleep in an animal and the appearance of an α rhythm in the electroencephalogram. Probably, the reciprocal principle of organizing ascending and descending projections is common to the entire system of the reticular formation.
What are the functions of the medulla oblongata
The functions performed by the human medulla oblongata are important for the body. Without this part of the brain, a person cannot live, since it controls the vital organs of the body - the lungs and heart muscle. Therefore, when this cerebral part of the central nervous system is injured, instant death occurs. Nuclei - accumulations of gray matter - are responsible for all this activity.
Table of functions of the nuclei of the medulla oblongata:
| Structure name | Tasks it performs |
| Nuclei of the V-XII pair of cranial nerves | Sensory, motor, autonomic functions of the hindbrain |
| Gray matter of the gracilis and cuneate fasciculus | Responsible for tactile and proprioceptive sensitivity |
| Olive kernels | Are part of the center of balance |
| Dorsal nucleus of the trapezius | Participate in the analysis of auditory information |
| Gray matter of the reticular formation | Depending on the situation, they activate or inhibit neurons of the spinal cord and the cerebral cortex of the central nervous system, forming autonomic centers |
| blue spot | The structures are capable of releasing a specific hormone that affects the excitability of individual zones of the cerebrum cortex |
Main functions
Thus, the functions of the human medulla oblongata are divided into several categories:
- Implementation and control of unconditioned reflexes (reflex function);
- Motor skills of protective and orienting actions (sensory function);
- Automaticity of the process of breathing and blood circulation (integrative function);
- Maintaining a balance of torso position and muscle tone (conductive function).
Sensory
The sensory department controls and processes the reception of incoming impulses that come to it from receptors throughout the human body. These receptive structures consist of 2 units:
- Sensoepithelial cells;
- Axons of afferent neurons.
There, for example, impulses from the respiratory center are processed - blood counts and the structure of the lung tissue are assessed. Based on the results of these data, blood saturation with oxygen and metabolic processes in the body subsequently occur. The sensory section of the medulla oblongata receives and transmits impulses from the digestive organs, hearing, vision, and tactile points for further processing.
The medulla oblongata perceives tactile information using the following structures:
- When facial skin receptors are activated, the neurons of the trigeminal nerve are excited;
- Taste receptors - glossopharyngeal nerve;
- Sound receptors – cochlear nerve;
- If stability is violated - olives.
Based on the data obtained, a further reaction of the body follows, which is expressed in the implementation of unconditional reflex reactions. They are also carried out by the medulla oblongata of the central nervous system. With insufficient oxygen saturation in the blood, a person begins to breathe deeply and look for a source of fresh air, thereby activating the motor function of the department.
Conductor
The oblongata section is the connecting link between the dorsal and terminal sections of the central nervous system. Afferent and efferent tracts pass through it: corticospinal, spinothalamic, rubrospinal.
With their help, information is transmitted to the higher structures of the cerebrum and returned back to the organs in the form of impulses. The olivospinal, vestibulospinal and reticulospinal tracts begin here. Through them, muscle tension and coordination of movements of the arms and legs are stabilized.
In the medulla oblongata, the corticoreticular tracts from the cortex, as well as the upward spreading fibers of proprioceptive susceptibility from the spinal cord, end. This feature allows you to feel parts of your own body relative to each other in space.
Integrative
The integrative function of the medulla oblongata is to carry out body reactions unrelated to reflex activity. Their appearance is associated with the work of terminal neurons, which are programmed to carry out regulatory processes in a certain order. They are implemented through other NS centers or directly.
The movements of the eyeballs when the head is tilted are realized on the basis of the joint activity of the nuclei of the vestibular and oculomotor systems of the brain. Some of the nerve cells of the reticular substance of the medulla oblongata control the tone and coordination of other nerve centers of the cerebrum.
Tasks
The main task of the medulla oblongata, based on the characteristics of its structure and the functions it performs, is to provide various reflexes. These include: protective, digestive, cardiovascular, tonic, and also responsible for lung ventilation and muscle tone.
How do protective reflexes work:
- when poison or poor-quality food enters the stomach, the gag reflex is triggered;
- when dust gets into the nasopharynx, sneezing occurs;
- mucus secreted in the nose protects the body from bacteria and viruses;
- coughing attacks clear the bronchi of mucus;
- tearing and blinking protects the eyes from foreign objects, and the corneas from drying out.
This part of the brain contains nerve centers responsible for many reflexes: digestion, breathing, muscle tone, sucking, blinking, cardiovascular, thermoregulation. This department takes part in processing information coming from all receptors in the body. It also controls movement and thought processes.
The respiratory control center works like this: neurons are excited under the influence of chemical stimuli. The center itself consists of several groups of neurons that belong to different parts of the medulla oblongata.
Vascular tone is controlled by the vasomotor center located in the medulla oblongata, which works together with the hypothalamus. Chewing occurs when the receptors in the oral cavity are irritated. In the medulla oblongata, salivation is regulated, thereby controlling the volume and composition of saliva.