What does the conductive part of the sense organs consist of?
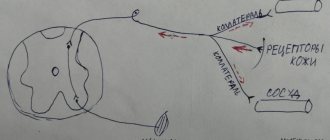
Conductor (conducting pathways - conductive part) is a sequence of neurons and their processes that transmit sensitive information. Usually it is three-neuron :
- The body of the first neuron is in the sensitive nodes of the CN (the I and II pairs of the CN have no sensitive nodes). The I and II pairs of CNs transmit impulses along the optic + olfactory nerves, they are false. Neurons are pseudounipolar, sensitive. But the VIII pair (spiral ganglion) has bipolar neurons. The sensory neuron has a process that reaches the receptor.
- The bodies of II neurons are in the sensory nuclei of the CN. The exception is pairs I and II (they do not contain sensitive nuclei). Axons of II neurons - through the trunk of the GM => decussation => bodies of III neurons.
- The bodies of III neurons are in the subcortical centers. Axons of III neurons go to the cortex. This is the last switching point + preliminary analysis takes place here. Some paths may close here.
What adaptive mechanisms are provided to the peripheral part of the organ of vision?
The peripheral part of the visual analyzer is the eye. Components of the eye: the eyeball and the auxiliary apparatus of the eye.
Eyeball
The eyeball is spherical in shape, consists of 3 shells and contains transparent media inside.
Eye shells
A) Fibrous membrane (outside):
- the anterior part is the cornea (transparent - does not contain blood vessels),
- The sclera is white and contains blood vessels.
B) The choroid is connective tissue and has more blood vessels than fibers. They nourish all shells. Includes three components (front to back):
- The iris is a colored disc with a hole in the center (the pupil). Coloring due to the pigment melanin. There are muscles that change the diameter of the pupil to regulate the flow of light. Only the pupil allows light to pass through.
- The ciliary body is a ring-shaped thickening where the cornea meets the sclera. There are two components: Muscles that provide accommodation.
- Ciliary processes - produce moisture in the chambers of the eye.
B) Retina - retina. Consists of two layers:
- pigment layer – located deeper, adjacent to the choroid;
- receptive layer - has many receptors.
The retina is homogeneous, except for two places:
- The blind spot is the optic disc;
- The macula is the point of best vision.
The retina contains receptors: rods and cones.
Rods - object vision (various shades of gray) - black and white vision. Available on the entire surface of the retina.
Cones – color vision. They are found in the posterior part of the retina, most of them in the macula. They work in the light (that's why it's black and white at dusk).
The retina is an outgrowth of the diencephalon, the other two layers are outgrowths of the ectoderm of the facial area.
Environments of the eye
1) Transparent media of the eye - freely transmit light. They include:
- Anterior chamber - between the cornea and iris,
- The posterior chamber is between the lens and the iris.
Fluid => posterior chamber => pupil => anterior chamber => iridocorneal angle => venous sinus of the sclera (Schlemm's canal). This is how fluid drains out. This fluid nourishes the transparent media (they do not contain blood vessels) and provides intraocular pressure .
Thus, the anterior and posterior chambers of the eye are filled with fluid (humidity of the eye chambers) that does not contain cells.
The fluid is formed by the ciliary bodies:
- The lens is a doubly convex lens, a gel-like substance. The main property is the ability to change curvature. It is surrounded by a capsule, the ligament of Zinn is attached to the edges (goes to the ciliary body). It flattens the lens.
- The vitreous humor fills most of the apple. channels with moisture for nutrition pass through it.
2) Refractive media of the eye (concentration of light rays on the retina):
- Cornea (strongest refraction),
- Liquid chambers,
- Lens (capable of changing refractive power), Vitreous body.
Accessory eye apparatus
- Muscles of the eye (6 muscles, innervated by III, IV, VI pairs of CN);
- Eyelids;
- Conjunctiva;
- Brows;
- Lacrimal apparatus (moisturizing and protection) – washes the eyeball with tears:
- lacrimal gland,
- A tear flowing through the conjunctival cavity
- tear ducts,
- tear sacs,
- Nasolacrimal duct.
Cortical ends (centers) of analyzers.
The doctrine of the cytoarchitectonics of the cerebral cortex corresponds to the teaching of I.P. Pavlova about the cortex as a system of cortical ends of analyzers. The analyzer, according to Pavlov, “is a complex nervous mechanism that begins with the external perceptive apparatus and ends in the brain.” The analyzer consists of three parts - the external perceptive apparatus (sensory organ), the conductive part (conducting tracts of the brain and spinal cord) and the final cortical end (center ) in the cerebral cortex of the telencephalon. According to Pavlov, the cortical end of the analyzer consists of a “core” and “scattered elements.”
The analyzer core is divided according to its structural and functional features into the central field of the nuclear zone and the peripheral field. In the first, finely differentiated sensations are formed, and in the second, more complex forms of reflection of the external world.
Scattered elements are those neurons that are located outside the nucleus and carry out simpler functions.
Based on morphological and experimental-physiological data in the cerebral cortex, the most important cortical ends of the analyzers (centers), which through interaction provide brain functions, have been identified.
The localization of the core analyzers is as follows:
Cortical end of the motor analyzer (precentral gyrus, precentral lobule, posterior section of the middle and inferior frontal gyri). The precentral gyrus and the anterior section of the pericentral lobule are part of the precentral region - the motor or motor zone of the cortex (cytoarchitectonic fields 4, 6). In the upper part of the precentral gyrus and the precentral lobule there are motor nuclei of the lower half of the body, and in the lower part - the upper half. The largest area of the entire zone is occupied by the centers of innervation of the hand, face, lips, tongue, and the smaller area is occupied by the centers of innervation of the muscles of the trunk and lower extremities. This area was previously considered to be only motor, but is now considered to be the region containing interneurons and motor neurons. Interneurons perceive stimuli from proprioceptors of bones, joints, muscles and tendons. The centers of the motor zone innervate the opposite part of the body. Dysfunction of the precentral gyrus leads to paralysis on the opposite side of the body.
The core of the motor analyzer for combined rotation of the head and eyes in the opposite direction, as well as the Motor nuclei of written speech - graphs related to voluntary movements associated with writing letters, numbers and other characters are localized in the posterior part of the middle frontal gyrus (field and on the border of the parietal and occipital lobe (field 19). The center of the graph is closely connected with field 40, located in the supramarginal gyrus. If this area is damaged, the patient cannot make the movements necessary to draw letters.
and on the border of the parietal and occipital lobe (field 19). The center of the graph is closely connected with field 40, located in the supramarginal gyrus. If this area is damaged, the patient cannot make the movements necessary to draw letters.
The premotor zone is located anterior to the motor areas of the cortex (fields 6 and 8). The processes of the cells of this zone are connected both with the nuclei of the anterior horns of the spinal cord, and with the subcortical nuclei, red nucleus, substantia nigra, etc.
The core of the motor analyzer of speech articulation (speech-motor analyzer) is located in the posterior part of the inferior frontal gyrus (field 44, 45, 45a). In field 44 - Broca's area, in right-handed people - in the left hemisphere, the analysis of irritations from the motor apparatus is carried out, through which syllables, words, and phrases are formed. This center was formed next to the projection area of the motor analyzer for the muscles of the lips, tongue, and larynx. When it is affected, a person is able to pronounce individual speech sounds, but he loses the ability to form words from these sounds (motor or motor aphasia). In case of damage to field 45, the following is observed: agrammatism - the patient loses the ability to compose words from words, to coordinate words into sentences.
The cortical end of the motor analyzer of complex coordinated movements in right-handers is located in the inferior parietal lobule (area 40) in the region of the supramarginal gyrus. If field 40 is affected, the patient, despite the absence of paralysis, loses the ability to use household items and loses production skills, which is called apraxia.
The cortical end of the skin analyzer of general sensitivity - temperature, pain, tactile, muscle-articular - is located in the postcentral gyrus (fields 1, 2, 3, 5). Damage to this analyzer results in loss of sensitivity. The sequence of locations of the centers and their territory corresponds to the motor zone of the cortex.
The cortical end of the auditory analyzer (field 41) is located in the middle part of the superior temporal gyrus.
The auditory analyzer of oral speech (control of one's own speech and perception of someone else's) is located in the posterior part of the superior temporal gyrus (field 42) (Wernicke's area_ when it is disrupted, a person hears speech, but does not understand it (sensory aphasia)
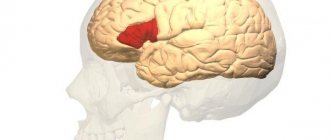
The cortical end of the visual analyzer (fields 17, 18, 19) occupies the edges of the calcarine groove (field 17); complete blindness occurs with bilateral damage to the nuclei of the visual analyzer. In cases of damage to fields 17 and 18, loss of visual memory is observed. If field 19 is damaged, a person loses the ability to navigate in a new environment.
The visual analyzer of written signs is located in the angular gyrus of the inferior parietal lobule (area 39s). If this field is damaged, the patient loses the ability to analyze written letters, that is, loses the ability to read (Alexia)
The cortical ends of the olfactory analyzer are located in the parahippocampal gyrus on the inferior surface of the temporal lobe and the hippocampus.
The cortical ends of the taste analyzer are in the lower part of the postcentral gyrus.
The cortical end of the stereognostic sense analyzer , the center of a particularly complex type of recognition of objects by touch, is located in the superior parietal lobule (field 7). When the parietal lobule is damaged, the patient cannot recognize an object by feeling it with the hand opposite the lesion - stereognosia. There are auditory gnosis - recognition of objects by sound (a bird - by voice, a car - by the noise of engines), visual gnosis - recognition of objects by sight, etc. Praxia and gnosis are functions of a higher order, the implementation of which is associated with both the first and with a second signaling system, which is a specific human function.
Any function is not localized in one specific field, but is only predominantly associated with it and spreads over a large area.
Speech is one of the phylogenetically new and most complexly localized functions of the cortex, associated with the second signaling system, according to I.P. Pavlova. Speech appeared in the course of human social development, as a result of labor activity. “...First, work, and then, along with it, articulate speech were the two most important stimuli, under the influence of which the monkey’s brain gradually turned into the human brain, which, for all its similarities with monkeys, far surpasses it in size and perfection” (K. Marx, F. Engels)
The function of speech is extremely complex. It cannot be localized in any part of the cortex; the entire cortex participates in its implementation, namely neurons with short processes located in its superficial layers. With the development of new experience, speech functions can move to other areas of the cortex, such as gesturing in the deaf and dumb, reading in the blind, writing with the foot in the armless. It is known that in the majority of right-handed people, speech functions, functions of recognition (gnosia), and purposeful action (praxia) are associated with certain cytoarchitectonic fields of the left hemisphere, while in left-handers it is the other way around.
Association zones of the cortex occupy the remaining significant part of the cortex; they are devoid of obvious specialization and are responsible for combining and processing information and programmed actions. The associative cortex forms the basis of higher processes, such as memory, learning, thinking, and speech.
There are no zones that give rise to thoughts. To make the most insignificant decision, the entire brain is involved, various processes occurring in different zones of the cortex and in the lower nerve centers come into play.
The cerebral cortex receives information, processes it and stores it in memory. In the process of adaptation (adaptation) of the body to the external environment, complex systems of self-regulation and stabilization were formed in the cortex, providing a certain level of function, self-learning systems with a memory code, control systems operating on the basis of a genetic code, taking into account age and ensuring an optimal level of control and functions in the body , comparison systems that provide a transition from one form of management to another.
Connections between the cortical ends of a particular analyzer with the peripheral parts (receptors) are carried out by a system of pathways of the brain and spinal cord and peripheral nerves extending from them (cranial and spinal nerves).
Subcortical nuclei. They are located in the white matter of the base of the telencephalon and form three paired clusters of gray matter: the striatum, the amygdala and the fence, which make up approximately 3% of the volume of the hemispheres.
The striatum consists of two nuclei: the caudate and the lentiform.
The caudate nucleus is located in the frontal lobe and is a formation in the form of an arc lying on top of the optic thalamus and the lenticular nucleus. It consists of a head, body and tail, which take part in the formation of the lateral part of the wall of the anterior horn of the lateral ventricle of the brain.
The lenticular nucleus is a large pyramidal-shaped accumulation of gray matter located lateral to the caudate nucleus. The lentiform nucleus is divided into three parts: the outer, dark-colored shell and two light medial stripes - the outer and inner segments of the globus pallidus.
The caudate and lenticular nuclei are separated from each other by a layer of white matter - part of the internal capsule. Another part of the internal capsule separates the lenticular nucleus from the underlying thalamus.
The striatum forms a striopallidal system, in which the phylogenetically more ancient structure is the globus pallidus, the pallidum. It is separated into an independent morpho-functional unit that performs a motor function. Thanks to connections with the red nucleus and the substantia nigra of the midbrain, the pallidum carries out movements of the torso and arms when walking - cross-coordination, a number of auxiliary movements when changing body positions, facial movements. Destruction of the globus pallidus causes muscle rigidity.
The caudate nucleus and putamen are younger structures of the striatum - the striatum, which does not directly have a motor function, but performs a controlling function in relation to the pallidum, somewhat inhibiting its influence.
When the caudate nucleus is damaged, a person experiences rhythmic involuntary movements of the limbs (Huntington's chorea), and when the putamen is degenerated, trembling of the limbs occurs (Parkinson's disease).
The fence is a relatively thin strip of gray matter located between the insular cortex, separated from it by white matter - the outer capsule and the shell, from which it is separated by the outer capsule. The fence is a complex formation, the connections of which have so far been poorly studied, and the functional significance is not clear.
The amygdala is a large nucleus located under the shell in the depths of the anterior temporal lobe, has a complex structure and consists of several nuclei that differ in cellular composition. The amygdala is a subcortical olfactory center and is part of the limbic system.
The subcortical nuclei of the telencephalon function in close relationship with the cerebral cortex, diencephalon and other parts of the brain, and take part in the formation of both conditioned and unconditioned reflexes.
Together with the red nucleus, the substantia nigra of the midbrain, the thalamus of the diencephalon, the subcortical nuclei form the extrapyramidal system, carrying out complex unconditioned reflex motor acts.
olfactory brain is the oldest part of the telencephalon, which arose in connection with olfactory receptors. It is divided into two sections: peripheral and central.
The peripheral part includes: the olfactory bulb, the olfactory tract, the olfactory triangle and the anterior perforated substance.
The central section includes: the vaulted gyrus, consisting of the cingulate gyrus, the isthmus and the parahippocampal gyrus, as well as the hippocampus - a peculiarly shaped formation located in the cavity of the inferior horn of the lateral ventricle and the dentate gyrus, which lies inside the hippocampus.
The limbic system (edge, edge) is so named because the cortical structures included in it are located on the edge of the neocortex and, as it were, border the brain stem. The limbic system includes both certain zones of the cortex (archipaleocortical and interstitial areas) and subcortical formations.
Of the cortical structures, these are: the hippocampus with the dentate gyrus (old cortex), the cingulate gyrus (the limbic cortex, which is interstitial), the olfactory cortex, the septum (ancient cortex).
From the subcortical structures: the mamillary body of the hypothalamus, the anterior nucleus of the thalamus, the amygdala complex, and the fornix.
In addition to numerous two-way connections between the structures of the limbic system, there are long paths in the form of closed circles along which excitation circulates. The greater limbic circle - the Peipets circle includes: the hippocampus, fornix, mamillary body, mastoid-thalamic fascicle (Vic d'Azir fascicle), anterior nucleus of the thalamus, cingulate cortex, hippocampus. Of the overlying structures, the limbic system has the closest connections with the frontal cortex. The limbic system directs its descending pathways to the reticular formation of the brain stem and to the hypothalamus.
Through the hypothalamic-pituitary system, it exercises control over the humoral system. The limbic system is characterized by special sensitivity and a special role in the functioning of hormones synthesized in the hypothalamus, oxytocin and vasopressin, secreted by the pituitary gland.
The main integral function of the limbic system is not only the olfactory function, but also the reactions of the so-called innate behavior (eating, sexual, searching and defensive). It carries out the synthesis of afferent stimuli, is important in the processes of emotional and motivational behavior, organizes and ensures the flow of vegetative, somatic and mental processes during emotional and motivational activity, carries out the perception and storage of emotionally significant information, the selection and implementation of adaptive forms of emotional behavior.
Thus, the functions of the hippocampus are associated with memory, learning, the formation of new behavior programs when conditions change, and the formation of emotional states. The hippocampus has extensive connections with the cerebral cortex and the hypothalamus of the diencephalon. In mentally ill patients, all layers of the hippocampus are affected.
At the same time, each structure included in the limbic system contributes to a single mechanism, having its own functional characteristics.
The anterior limbic cortex provides emotional expressiveness to speech.
The cingulate gyrus takes part in reactions of alertness, awakening, and emotional activity. It is connected by fibers to the reticular formation and the autonomic nervous system.
The amygdala complex is responsible for feeding and defensive behavior; stimulation of the amygdala causes aggressive behavior.
The septum takes part in retraining, reduces aggressiveness and fear.
Mamillary bodies play an important role in the development of spatial skills.
Anterior to the fornix, in its various sections, are the centers of pleasure and pain.
The lateral ventricles are the cavities of the hemispheres of the telencephalon. Each ventricle has a central part adjacent to the superior surface of the optic thalamus in the parietal lobe and three horns extending from it.
The anterior horn extends into the frontal lobe, the posterior horn into the occipital lobe, and the inferior horn into the depths of the temporal lobe. In the lower horn there is an elevation of the inner and partially lower wall - the hippocampus. The medial wall of each anterior horn is a thin transparent plate. The right and left plates form a common transparent septum between the anterior horns.
The lateral ventricles, like all ventricles of the brain, are filled with cerebral fluid. Through the interventricular foramina, which are located in front of the visual thalamus, the lateral ventricles communicate with the third ventricle of the diencephalon. Most of the walls of the lateral ventricles are formed by the white matter of the telencephalon hemispheres.
White matter of the telencephalon. It is formed by fibers of conductive tracts, which are grouped into three systems: associative or combinational, commissural or commissural and projection.
Association fibers of the telencephalon connect different areas of the cortex within the same hemisphere. They are divided into short fibers, lying superficially and arcuately, connecting the cortex of two adjacent gyri, and long fibers, lying deeper and connecting areas of the cortex distant from each other. These include:
1) The belt, which can be traced from the anterior perforated substance to the hippocampal gyrus and connects the gyral cortex of the medial part of the surface of the hemisphere, belongs to the olfactory brain.
2) The inferior longitudinal fasciculus connects the occipital lobe with the temporal lobe and runs along the outer wall of the posterior and inferior horn of the lateral ventricle.
3) The superior longitudinal fasciculus connects the frontal, parietal and temporal lobes.
4) The uncinate fasciculus connects the rectum and orbital gyri of the frontal lobe with the temporal lobe.
Commissural nerve pathways connect the cortical areas of both hemispheres. They form the following commissures or commissures:
1) The corpus callosum is the largest commissure that connects various areas of the neocortex of both hemispheres. In humans it is much greater than in animals. In the corpus callosum, there is an anterior end curved downwards (beaked) - the knee of the corpus callosum, a middle part - the trunk of the corpus callosum and a thickened posterior end - the splenium of the corpus callosum. The entire surface of the corpus callosum is covered with a thin layer of gray matter - the gray vesture.
In women, more fibers pass through a certain area of the corpus callosum than in men. Thus, interhemispheric connections in women are more numerous, and therefore they are better able to integrate information available in both hemispheres, which explains gender differences in behavior.
2) The anterior callosal commissure is located behind the beak of the corpus callosum and consists of two bundles; one connects the anterior perforated substance, and the other connects the gyri of the temporal lobe, mainly the hippocampal gyrus.
3) The vault commissure connects the central parts of two arcuate bundles of nerve fibers, which form the vault located under the corpus callosum. The vault is divided into a central part - the pillars of the vault and the legs of the vault. The columns of the fornix connect a triangular plate - the commissure of the fornix, the posterior part of which is fused with the lower surface of the corpus callosum. The columns of the fornix, curving posteriorly, enter the hypothalamus and end in the mamillary bodies.
Projection pathways connect the cerebral cortex with the nuclei of the brain stem and spinal cord. There are: efferent - descending motor pathways that conduct nerve impulses from the cells of the motor areas of the cortex to the subcortical nuclei, motor nuclei of the brain stem and spinal cord. Thanks to these pathways, the motor centers of the cerebral cortex are projected to the periphery. Afferent - ascending sensory pathways are processes of cells of the spinal ganglia and ganglia of the cranial nerves - these are the first neurons of the sensory pathways that end on the switching nuclei of the spinal cord or medulla oblongata, where the second neurons of the sensory pathways are located, going as part of the medial loop to the ventral nuclei of the thalamus. In these nuclei lie the third neurons of the sensory pathways, the processes of which go to the corresponding nuclear centers of the cortex.
Both sensory and motor pathways form in the substance of the cerebral hemispheres a system of radiating fascicles - the corona radiata, which gathers into a compact and powerful bundle - the internal capsule, which is located between the caudate and lenticular nuclei, on the one hand, and the thalamus, on the other hand. It distinguishes between the front leg, the knee and the back leg.
The pathways of the brain are the spinal cords.
The membranes of the brain. The brain, like the spinal cord, is covered with three membranes - dura mater, arachnoid membrane and vascular membrane.
The dura mater of the brain differs from that of the spinal cord in that it is fused to the inner surface of the skull bones and there is no epidural space. The dura mater forms channels for the outflow of venous blood from the brain - the sinuses of the dura mater and gives rise to processes that provide fixation of the brain - these are the falx cerebri (between the right and left hemispheres of the brain), the tentorium cerebellum (between the occipital lobes and the cerebellum) and the diaphragm sella (above sella turcica, in which the pituitary gland is located). In the places where the processes depart, the dura mater is stratified, forming sinuses, where venous blood of the brain, dura mater, and skull bones flows into the system of external veins through the graduates.
The arachnoid membrane of the brain is located under the dura mater and covers the brain without entering its grooves, spreading over them in the form of bridges. On its surface there are outgrowths - Pachionian granulations, which have complex functions. Between the arachnoid and choroid, a subarachnoid space is formed, well defined in the cisterns that form between the cerebellum and medulla oblongata, between the cerebral peduncles, in the region of the lateral sulcus. The subarachnoid space of the brain communicates with those of the spinal cord and the fourth ventricle and is filled with circulating cerebral fluid.
The choroid of the brain consists of 2 plates, between which arteries and veins are located. It is closely fused with the substance of the brain, enters all the cracks and grooves and participates in the formation of choroid plexuses, rich in blood vessels. Penetrating into the ventricles of the brain, the choroid produces cerebral fluid, thanks to its choroid plexuses.
Lymphatic vessels were not found in the meninges.
The innervation of the meninges is carried out by the V, X, XII pairs of cranial nerves and the sympathetic nerve plexus of the internal carotid and vertebral arteries.
LECTURE No. 5
How many neurons does the visual pathway consist of? Where are the subcortical and cortical vision centers located?
The visual pathway consists of the bodies of 4 neurons and their processes.
- Axons of I and II neurons (short) together with their bodies are located in the retina.
- Axons of III neurons form the optic nerve: Eyeball => optic canal => cranial cavity => partial optic chiasm (between two nerves) => an optic tract is formed at the base of the skull, heading to the subcortical centers of vision.
- The bodies of IV neurons are in the subcortical centers of vision: lateral geniculate bodies,
- superior colliculus,
- the posterior nuclei of the thalamus are in the thalamic cushion.
What is a sound-conducting apparatus?
The sound follows the following chain:
- Auricle,
- External auditory canal,
- Eardrum,
- auditory ossicles,
- Perilymph of the scala vestibule,
- drum ladder,
- Endolymph of the cochlear duct.
The sound-receiving part is the organ of Corti.
Its functions:
- Resonance,
- Fluctuations in the endo- and perilymph => excitation of nerve cells,
- Located on the basement membrane along the entire length of the cochlear duct.
Cortical centers
Finite brain
The telencephalon is a derivative of the forebrain and is represented by two hemispheres of the cerebrum. In each hemisphere there are: 1) a mantle formed from the dorsal wall of the brain bladder; 2) olfactory brain ;
3) basal ganglia, developing from its ventral section. Inside each hemisphere there is a cavity - the lateral ventricle communicating with the third ventricle.
The outer layer of the cloak is the cortex, under which lies the white matter, which makes up the largest part of the hemisphere.
BRAIN CORTICAL HEMISPHERES
The cerebral cortex is a layer of gray matter, the thickness of which varies in different parts and averages 2-3 mm. The surface of the cortex has a complex topography, characterized by numerous grooves and elevations located between them - convolutions .
The gyri differ from each other in shape and size, but the gyri of the same name on the cerebral cortex of different people are fundamentally similar and are localized in certain places.
In each hemisphere of the cerebrum, there are dorsolateral, medial and inferior surfaces. The dorsolateral surface of the hemispheres is the most extensive, has a convex shape, faces upward and laterally, and borders the medial surface with a clearly defined edge. The flat medial surface faces the longitudinal fissure of the brain, in the middle part it is connected by the corpus callosum with the same surface of the other hemisphere. The lower surface is flattened in the anterior section and concave in the posterior section. Three main sulci divide each hemisphere into four lobes.
- The lateral sulcus begins on the lower surface of the hemisphere in the form of the lateral (Sylvian) fossa of the cerebrum, runs along the lateral side upward and backward. It is the anterior superior border of the temporal lobe, ,
and separates the frontal and temporal lobes.
- The central sulcus runs frontally along the dorsolateral surface of the hemisphere, starting from its upper edge. Usually it passes to its medial side and the lower part does not reach the Sylvian fissure. It divides the upper part of the hemisphere into the anterior (smaller) section - the frontal lobe, the posterior (larger) section, including the parietal lobe, and the occipital lobe. A characteristic feature of the central sulcus is its continuity throughout its entire length.
- The parieto-occipital sulcus, sulcus
parietooccipitalis ,
is located in the posterior part of the brain on the medial surface of the hemisphere, continuing slightly to the dorsolateral surface. This groove is the boundary between the parietal and occipital lobes.
Let us consider the relationship between the lobes of the cerebral hemisphere. The frontal lobe occupies the dorsolateral surface of the hemisphere anterior to the central sulcus; the lower surface is anterior to the lateral groove. On the medial surface there is no real border between the frontal and parietal lobes; here they are separated by an imaginary continuation of the central sulcus.
The temporal lobe occupies the dorsolateral surface inferior to the lateral sulcus and the inferior surface of the hemisphere posterior to the lateral (Sylvian) fossa of the cerebrum. On the medial surface it is located below the brain stem.
The parietal lobe lies in the center of the brain. On the dorsolateral surface, it belongs to the area of the hemisphere between the central sulcus in front, the lateral sulcus below and the imaginary continuation of the parieto-occipital sulcus. On the medial surface of the hemisphere, the parietal lobe occupies the area between the parieto-occipital sulcus, an imaginary continuation of the central sulcus in front and the corpus callosum below.
The occipital lobe is clearly delimited from the parietal lobe only on the medial surface by the parieto-occipital sulcus. On the dorsolateral and inferior surfaces of the hemisphere, its border is drawn by imaginary lines that are continuations of the indicated groove.
In addition to the four lobes described, there is also an insula.
It lies deep in the lateral sulcus and is visible only when moving apart the convolutions that limit this sulcus.
RELIEF OF THE DORSO-LATERAL SURFACE OF THE HEMISPHERES
Frontal lobe.
On the dorsolateral surface, the superior precentral sulcus and the inferior precentral sulcus pass in front of the central sulcus.
Sometimes they merge into one precentral sulcus .
From these two grooves, two frontal grooves originate, moving forward: superior and inferior (Fig. 49).
These four grooves divide the surface of the frontal lobe into the following gyri. Anterior to the central sulcus is the precentral gyrus. In the remaining area, three frontal gyri stand out: the superior frontal gyrus, located above the no
the upper edge of the hemisphere;
middle frontal gyrus, lies between the superior and inferior frontal sulci; inferior frontal gyrus, located between and .
Parietal lobe .
On the dorsolateral surface, the postcentral sulcus runs parallel to the central sulcus
.
From it begins in the sagittal direction the long intraparietal
furrow.
These two grooves divide the surface of the parietal lobe into three sections.
Between the central and postcentral sulci is the postcentral gyrus .
Upwards it continues to the medial surface of the hemisphere.
The area of cortex located above the intraparietal sulcus is
called the superior parietal lobule
.
The underlying area is the inferior parietal lobule
.
It contains two very important gyri: supramarginal, which closes the end of the lateral sulcus
,
and angular
,
which surrounds the posterior end of the superior temporal gyrus
.
Occipital lobe .
The smallest of all shares.
On its dorsolateral surface, the grooves vary greatly. Here, a distinction is made between the superior occipital sulcus and
the lateral occipital sulcus
.
In accordance with this, superior and lateral occipital gyri are distinguished
.
Temporal lobe .
On the dorsolateral surface in the anteroposterior direction there pass the superior temporal sulcus
,
which with its posterior end extends into the region of the parietal lobe, and the highly variable middle temporal sulcus
.
The last sulcus is located closer to the lower edge of the temporal lobe.
The inferior temporal sulcus is
located already on the basal surface.
On the dorsolateral surface of the temporal lobe are the superior temporal gyrus and the middle temporal gyrus..
They are separated from each other by the superior temporal sulcus.
The inferior temporal gyrus runs along the lower edge of the hemisphere, bounded
by the middle and inferior temporal sulci.
Island).
The insula is clearly visible only when moving apart the edges of the lateral sulcus or after removing the overhanging tegmental (opercular) sections of the frontal, parietal and temporal lobes, which limit the lateral sulcus, at the bottom of which it is located.
The insula bears some resemblance to a cone, the base of which is surrounded by a deep circular insula groove .
Its surface is divided by the central groove of the insula into anterior and posterior lobes.
The posterior lobe usually consists of only one long convolution of the insula, the anterior lobe contains several short convolutions of the insula .
RELIEF OF THE MEDIAL SURFACE OF THE HEMISPHERES
Furrows on the medial surface of the cerebral hemispheres
(Fig. 50).
onto
the medial surface of the cerebral hemisphere.
The main sulcus is the sulcus of the corpus callosum, which surrounds the corpus callosum on its convex side, continuing into the hippocampal sulcus .
Approximately halfway between the sulcus of the corpus callosum and the upper edge of the hemisphere, the cingulate sulcus is located
.
She turns to
the upper edge of the hemisphere with its posterior end - the marginal branch, and extends slightly onto the dorsolateral surface, posterior to the central sulcus. In front of the marginal branch, approximately above the middle of the corpus callosum, the cingulate gyrus gives off the paracentral sulcus upward .
The direct continuation of the cingulate sulcus is the subparietal sulcus
.
Below the posterior end of the corpus callosum, two grooves begin with a common trunk, diverging to the edge of the hemisphere: the already described parieto-occipital groove, and the calcarine groove
.
Near the occipital pole, a collateral groove begins on the lower surface of the hemisphere, heading anteriorly.
Its continuation in the anterior part of the temporal lobe is the nasal sulcus .
Lateral to the collateral sulcus is the inferior temporal sulcus
.
Convolutions of the medial surface of the cerebral hemispheres.
The part of the medial surface lying above the cingulate gyrus belongs to the frontal lobe.
This is the superior frontal gyrus extending here. Posteriorly, it reaches the level of the projection of the upper end of the central sulcus. Within the parietal lobe there is a pericentral lobule, which below reaches the subparietal sulcus .
The circumcentral lobule connects the parietal lobe with the frontal lobe on the medial surface
.
Between
purs marginalis sulci cinguli
- in front,
sulcus parietooccipitalis
- behind and
sulcus
subparietalis
— the precuneus lies below,
.
Between
the sulcusparietooccipitalis
and
sulcus calcarinus
(already in the occipital lobe) there is a wedge,
cutieus .
On the medial surface of the same lobe is the lingual gyrus, lying between
the sulcus carcarinus
and
sulcus collateralis .
Below the latter is the medial occipitotemporal gyrus
.
Within the temporal lobe, on the medial surface of the hemispheres, directly below the cerebral peduncles, there is the parahippocampal gyrus, which ends anteriorly in the uncus .
The parahippocampal gyrus and uncus are separated from the cerebral peduncles by the hippocampal sulcus
.
Below the parahippocampal gyrus lies the lateral occipitotemporal gyrus
.
These gyri are separated posteriorly by the collateral groove and in front by the nasal groove
.
The inferior temporal gyrus runs along the lowest edge of the medial surface of the temporal lobe, which is separated by the groove of the same name from the lateral occipitotemporal gyrus.
The gyri, ring-shaped bordering the corpus callosum and the cerebral peduncles, extending from the frontal lobe to the temporal lobe, as a whole make up the vaulted gyrus,
which is distinguished as the limbic lobe,
lobus / imbicus .
It consists of two parts: the cingulate gyrus,
gyrus cinguli ,
and the parahippocampal gyrus,
gyrus parahippocampalis ,
connected to each other behind the splenium of the corpus callosum, isthmus,
isthmus gyri cinguli .
The cingulate gyrus lies between the sulcus of the corpus callosum, on the one hand, and the cingulate sulcus and the subparietal sulcus, on the other. The parahippocampal gyrus, as already noted, is bounded above by the hippocampal sulcus, sulcus
hippocampi ,
and below by the anterior end of the collateral and nasal sulci.
RELIEF OF THE LOWER SURFACE OF THE HEMISPHERES
On the lower (basal) surface of the frontal lobe there is an olfactory groove, sulcus
olfactorius ,
running parallel to the longitudinal fissure of the brain, and more laterally - orbital grooves,
sulci orbitales .
Between these grooves there are gyri of variable shape: straight gyrus,
gyrus rectus ,
which is bounded by
sulcus olfactorius
and
fissura longitudinalis cerebri ,
and orbital gyri,
gyri orbitales ,
lying lateral to the olfactory groove (Fig. 51).
Within the temporal and occipital lobes there is no clear boundary between the medial and inferior surfaces. They gradually transform into each other. In this regard, the grooves and convolutions located on the medial surface of the hemispheres in the lower parts of the occipital and temporal lobes are also visible on the lower surface of the hemispheres. In particular, within the occipital lobe is the medial occipitotemporal gyrus. Within the temporal lobe lie the parahippocampal, lateral occipitotemporal and inferior temporal gyri. The sequence of location of the named convolutions is considered in the lateral direction. The grooves separating these convolutions were named earlier.
The given description of the grooves and convolutions of the cerebral cortex can be considered schematic, since individual variants of their architectonics are quite common.
STRUCTURE OF THE CEREBRAL HEMISPHERES CORTEX
The cerebral cortex, cortex
cerebri ,
constitutes the most important part of the brain, being the material substrate of higher nervous activity and the main regulator of all vital functions of the body. The cortex carries out analysis and synthesis of incoming stimuli from the internal environment of the body and
from the surrounding external environment. Consequently, the highest forms of reflection of the external world and conscious human activity are associated with the cerebral cortex.
The formation of the cortex is one of the most progressive acquisitions in the evolution of the vertebrate brain. Phylogenetic approaches to the study of this issue show that the cortex developed gradually. Thus, in fish and amphibians, the cerebral cortex performed only the function of an analyzer of olfactory stimuli. Reptiles develop a neocortex that analyzes visual information. In mammals, the neocortex already occupies most of the cerebral hemispheres. It analyzes impulses of all types of sensitivity and develops higher motor centers. In higher mammals, primates and humans, the rapid development of the neocortex leads to the formation of grooves and convolutions, due to which the surface of the cerebral hemispheres increases.
When considering the human cerebral cortex from a phylogenetic perspective, one can distinguish ancient, old and new cortex. To the ancient cortex, paleocortex ,
refers to a small area located on the ventral surface of the frontal lobe near the olfactory bulb.
The old cortex, archicortex ,
includes the hippocampus, or Ammon's horn, which is located in the cavity of the lateral ventricles of the telencephalon.
The displacement of the old cortex to the indicated location occurred as a result of significant growth of the new cortex, neocortex .
The neocortex in humans accounts for 95.4% of the entire surface of the cerebral hemisphere. In the human embryo, already in the 5th month of intrauterine development, grooves begin to form on the cerebral cortex. The lateral sulcus is formed first, followed by the central, parieto-occipital, calcarine and hippocampal sulci. From 7 months, the process of the appearance of furrows accelerates, and secondary furrows develop. By the time a child is born, the relief of the hemispheres is mainly formed. After birth, the formation of tertiary grooves is noted, which determine the individual characteristics of the relief of the hemispheres.
In an adult, due to the final formation of the sulci, the area of the cerebral hemispheres averages 1550 cm2:
The human neocortex, in comparison with the neocortex of other mammals, differs, in addition to its size, in its high differentiation. Its thickness is not the same everywhere. On average equal to 2-3 mm, it varies not only in different parts of the hemisphere, but also in different parts of the convolutions. It reaches its greatest development in the upper parts of the precentral and postcentral gyri, as well as in the paracentral lobule. At the tops of the gyri, the cortex is usually thicker than at the depths of the grooves. In general, the cortex accounts for 44% of the total volume of the hemispheres. The number of neurocytes in the cerebral cortex is on average 15 billion. However, in volume they are equal to only 1:27 of the volume of the entire cortex, 26 parts account for glial elements that provide supporting, protective and trophic functions in relation to neurocytes.
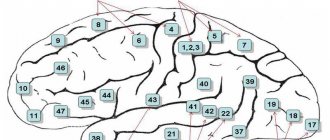
The founder of studies of the cellular composition of the cerebral cortex, features of the structure and distribution of nerve cells (cytoarchitecture of the cortex) is V. A. Betz. Subsequently, Brodman identified 52 fields in the cerebral cortex, designating each of them with a specific number. The same numbering of fields is preserved in the cytoarchitectonic
This is a map compiled by the Brain Institute of Russia, but in it a number of fields are divided into zones designated by letters of the Latin alphabet (Fig. 52).
Nerve cells are distributed unequally in different parts of the cerebral cortex. At the same time, it was discovered that nerve cells homogeneous in their structural characteristics are grouped into separate layers, the number of which in the mature cortex varies from 5 to 8. A significant part of the cerebral hemispheres has a six-layer type of cortical structure. In some areas, for example, in the precentral gyrus, gyms
precentralis ,
a reduction of the fourth layer usually occurs; in
the area striata
of the occipital lobe, on the contrary, it splits into three new ones.
The outermost layer is the molecular plate, lamina
molecularis ,
contains a small number of small nerve cells and is composed mainly of a dense plexus of nerve fibers lying parallel to the surface of the convolutions.
The second layer is the outer granular plate, laminagranu / aris
externa ,
which contains a large number of small, polygonal or round nerve cells.
20/
The third layer is the outer pyramidal plate, lamina
pyramidalis externa ,
consists of the same small cells as the second layer.
The fourth layer is called the internal granular plate, lamina
granularis interna .
The fifth layer is the layer of large pyramidal cells or ganglion cells, lamina
ganglionaris ,
represented by the internal pyramidal plate,
lamina pvramidalis interna .
Along with fairly large pyramidal cells, it also contains the so-called Betz giant pyramidal cells, which are found only in certain areas of the cortex: in the anterior central gyrus (mainly in its upper section) and in the paracentral lobule of the medial surface of the hemisphere. Pyramidal cells with their apex face the surface of the brain; the base from which the axon departs is to the white matter. The fifth layer gives rise to the efferent (descending) corticospinal and corticonuclear tracts.
The last layer, lying on the border of the white matter, is polymorphic, lamina
multiformis ,
which, as its name shows, contains cellular elements of the most diverse shapes (triangular, polygonal, oval, spindle-shaped).
It is customary to combine the three outer layers under the name of the main outer zone, and the three inner layers under the name of the main inner zone. The functional significance of the plates is determined by their cellular composition and interneuronal connections. The molecular plate ends with fibers from other layers of the cortex and from the opposite hemisphere. There is an opinion that neurons of the molecular plate are directly related to memory processes. The external granular and external pyramidal plates mainly contain associative neurons that carry out intracortical connections. They enable analytical thought processes. These plates are phylogenetically the youngest; they are highly developed in the cerebral cortex of humans.
The internal granular plate is the main afferent layer of the cortex. On the neurons of this plate, projection nerve fibers coming from the nuclei of the thalamus and geniculate bodies end. The efferent projection fibers of the cortex begin from the pyramidal cells of the internal pyramidal plate. The lamina multiforme contains functionally heterogeneous neurons. Associative and commissural fibers originate from them.
Currently, data have been obtained on the structural and functional relationship of cells in various layers of the cerebral cortex. In this regard, the concept of cortical columns, or modules, was introduced. Each cortical column is a vertically oriented row of neurons running through all layers of the cortex. The module has its own input and output and is designed to process incoming information. The number of neurons in the cortical columns is constant and is 110 in most fields, only in the visual fields it increases to 300-500. The cortical columns are surrounded by radially located arterioles and nerve fibers, i.e. have certain boundaries.
The fields of the cerebral cortex are also characterized by the specificity of myeloarchitecture: radial and tangential nerve fibers of the cortex are distinguished. Radial fibers enter the cortex from the white matter of the hemispheres or, conversely, are directed into it from the cortex. Tangential fibers are located parallel to the layers of the cortex and form plexuses (strips). The fibers originating in the strips are connected between the
fight neurons of neighboring cortical columns. The number of stripes in different fields of the cortex is not the same. There are especially many of them in the visual fields of the cortex. Based on the characteristics of myeloarchitectonics, Vogt identifies more than 100 fields in the cerebral cortex. Finally, the distribution of glia in different areas of the cortex—glioarchitecture—has significant differences.
DYNAMIC LOCALIZATION OF FUNCTIONS IN THE HEMISPHERE CORTEX OF THE BRAIN
Based on numerous clinical, pathological, electrophysiological and morphological studies, the functional significance of various areas of the cerebral cortex has been clearly established.
Areas of the cerebral cortex that have characteristic cytoarchitectonics and nerve connections involved in performing certain functions are nerve centers. Damage to such areas of the cortex manifests itself in the loss of their inherent functions. The nerve centers of the cloak can be divided into projection and associative.
Projection centers are areas of the cerebral cortex, representing the cortical part of the analyzer, which have a direct morphofunctional connection through afferent or efferent nerve pathways with neurons of the subcortical centers.
Associative centers are areas of the cerebral cortex that do not have a direct connection with subcortical formations, but are connected by a temporary two-way connection with projection centers. Associative centers play a primary role in the implementation of higher nervous activity.
At present, the dynamic localization of some functions of the cerebral cortex has been clarified quite accurately.
Areas of the cerebral cortex that are not projection or associative centers are involved in inter-analyzer integrative brain activity.
Projection nerve centers
The cerebral cortex develops both in humans and in higher vertebrates. They begin to function immediately after birth. The formation of these centers is completed much earlier than associative ones. The following projection centers are clinically important.
Projection center
of general sensitivity
(tactile, pain, temperature and conscious proprioceptive).
This nerve center is also called the skin analyzer of general sensitivity. It is localized in the cortex of the postcentral gyrus (fields 1, 2, 3). It ends with the fibers that run as part of the thalamo-cortical pathway, tr . thalamocorticalis .
Each area of the opposite half of the body has a distinct projection at the cortical end of the skin analyzer (somatotopic projection). In the upper part of the postcentral gyrus the lower limb and torso are projected, in the middle - the upper limb and in the lower - the head (Penfield's sensory homunculus). The size of the projection zones of the somatosensory cortex is directly proportional to the number of receptors located in the skin. This explains the presence of the largest
somatosensory areas corresponding to the face and hand. Damage to the postcentral gyrus causes loss of tactile, pain, temperature sensitivity and muscle-articular sensation on the opposite half of the body.
Projection center of motor functions (kinesthetic center), or
motor analyzer.
This center is located in the motor cortex, which includes the precentral gyrus and paracentral lobule (fields 4, 6).
In the 3rd-4th layers of the cortex of the motor analyzer, the fibers running as part of the thalamo-cortical pathway end. Here the analysis of proprioceptive (kinesthetic) stimuli is carried out. In the fifth layer of the cortex there is the nucleus of the motor analyzer, from the neurocytes of which the corticospinal and corticonuclear tracts, tr . corticospinalis
and
tr . corticonuclearis .
The precentral gyrus also has a clear somatotopic localization of motor functions. Muscles that perform complex and finely differentiated movements have a large projection area in the cortex of the precentral gyrus. The largest area is occupied by the projection of the muscles of the tongue, face and hand, the smallest by the projection of the muscles of the trunk and lower extremities. The somatotopic projection to the precentral gyrus is called the “Penfield motor homunculus.” The human body is projected on the gyrus “upside down”, and the projection is carried out on the cortex of the opposite hemisphere.
Afferent fibers ending in the sensitive layers of the cortex of the motor analyzer initially pass as part of the Gaulle, Burdach and nuclear-thalamic tracts, conducting conscious muscle-joint feeling and partially tactile impulses. Damage to the precentral gyrus (due to hemorrhages, injuries) leads to impaired perception of proprioceptive stimuli from skeletal muscles, ligaments, joints and periosteum. The resulting central paralysis is associated with damage to the neurons of the motor cortex, the axons of which end on the neurons of the motor nuclei of the cranial nerves or on the neurons of the own nuclei of the anterior horns of the spinal cord. The corticospinal and corticonuclear tracts conduct impulses that provide conscious movements and have an inhibitory effect on the segmental apparatus of the brain stem and spinal cord. The cortical center of the motor analyzer, through a system of associative fibers, has numerous connections with various cortical sensory centers (the center of general sensitivity, the center of vision, hearing, vestibular functions, etc.). These connections are necessary to perform integrative functions when performing voluntary movements.
Projection center of the body diagram.
This center is located in the parietal lobe in the region of the intraparietal sulcus (area 40s). It presents somatotopic projections of all parts of the body. The center of the body circuit receives impulses primarily from conscious proprioceptive sensitivity. The main functional purpose of this projection center is to determine the position of the body and its individual parts in space and assess muscle tone. When the superior parietal lobule is damaged, there is a violation of such functions as recognition of parts of one’s own body, sensation of extra limbs, and disturbances in determining the position of individual parts of the body in space.